I want to relay here a comment from Tom Clark, Center for Naturalism, to my Jan. 4 post on Overbye's NY Times free will article, so that more of you are likely to see the link it includes.
"There were some suggestions in Overbye's article that questioning free will is a *good* thing. Einstein said seeing we don't have free will keeps us humble, and psychologist Dan Wegner suggested it would allow us to understand evil and prompt us to reform people instead of merely paying them back."
"There's a roundup of recent articles on free will, including Overbye's, at http://www.naturalism.org/roundup.htm."
Showing posts with label self. Show all posts
Showing posts with label self. Show all posts
Thursday, January 18, 2007
Thursday, January 04, 2007
The Free Will debate...
Any of you who have read my "I-Illusion" piece or followed this blog will know that I have a continuing interest in the issue of free will. The science section in the Jan. 2 issue of The New York Times has a beautifully written essay by Dennis Overbye - "Free Will: Now You Have It, Now You Don't" - which gives the views of Dennett, Wegner, Libet, Silberstein and others. I'm tempted to give you huge chunks of the article, but will retrain myself to just a few clips:
Overbye: A bevy of experiments in recent years suggest that the conscious mind is like a monkey riding a tiger of subconscious decisions and actions in progress, frantically making up stories about being in control.
Silberstein: If people freak at evolution, etc., how much more will they freak if scientists and philosophers tell them they are nothing more than sophisticated meat machines, and is that conclusion now clearly warranted or is it premature?
Dennett: When we consider whether free will is an illusion or reality, we are looking into an abyss. What seems to confront us is a plunge into nihilism and despair.
Overbye: Dennett...is one of many who have tried to redefine free will in a way that involves no escape from the materialist world while still offering enough autonomy for moral responsibility, which seems to be what everyone cares about. ... Dennett argues, it is precisely our immersion in causality and the material world that frees us. Evolution, history and culture, he explains, have endowed us with feedback systems that give us the unique ability to reflect and think things over and to imagine the future. Free will and determinism can co-exist.
Dennett: All the varieties of free will worth having, we have...We have the power of imagination, to see and imagine futures...That’s what makes us moral agents...You don’t need a miracle to have responsibility.
Overbye also reviews the idea of freedom as a possible emergent phenomena that grows naturally in accordance with the laws of physics - like stock markets, brains, or the rules of democracy - that play by new rules once they are here.
Overbye: A bevy of experiments in recent years suggest that the conscious mind is like a monkey riding a tiger of subconscious decisions and actions in progress, frantically making up stories about being in control.
Silberstein: If people freak at evolution, etc., how much more will they freak if scientists and philosophers tell them they are nothing more than sophisticated meat machines, and is that conclusion now clearly warranted or is it premature?
Dennett: When we consider whether free will is an illusion or reality, we are looking into an abyss. What seems to confront us is a plunge into nihilism and despair.
Overbye: Dennett...is one of many who have tried to redefine free will in a way that involves no escape from the materialist world while still offering enough autonomy for moral responsibility, which seems to be what everyone cares about. ... Dennett argues, it is precisely our immersion in causality and the material world that frees us. Evolution, history and culture, he explains, have endowed us with feedback systems that give us the unique ability to reflect and think things over and to imagine the future. Free will and determinism can co-exist.
Dennett: All the varieties of free will worth having, we have...We have the power of imagination, to see and imagine futures...That’s what makes us moral agents...You don’t need a miracle to have responsibility.
Overbye also reviews the idea of freedom as a possible emergent phenomena that grows naturally in accordance with the laws of physics - like stock markets, brains, or the rules of democracy - that play by new rules once they are here.
Wednesday, January 03, 2007
The distinction between sincerity and authenticity
I want to pass on clips from an essay by Orlando Patterson in the Dec. 26 New York Times. He cites Lionel Trilling, the cultural critic, as having in the 1970s "encouraged us to take seriously the distinction between sincerity and authenticity. Sincerity, he said, requires us to act and really be the way that we present ourselves to others. Authenticity involves finding and expressing the true inner self and judging all relationships in terms of it."
Patterson suggests that "Authenticity now dominates our way of viewing ourselves and our relationships, with baleful consequences. Within sensitive individuals it breeds doubt; between people it promotes distrust; within groups it enhances group-think in the endless quest to be one with the group’s true soul; and between groups it is the inner source of identity politics...the primacy of the self has penetrated major areas of government: emotivist arguments trump reasoned discourse in Congressional hearings and criminal justice; and in public education."
"Social scientists and pollsters routinely belittle results showing growing tolerance; they argue that Americans have simply learned how to conceal their deeply...Harvard social psychologist Mahzarin Banaji and her collaborators claim to have evidence, based on more than three million self-administered Web-based tests, that nearly all of us are authentically bigoted to the core with hidden “implicit prejudices” — about race, gender, age, homosexuality and appearance — that we deny, sometimes with consciously tolerant views ingrained prejudices."
"I couldn’t care less whether my neighbors and co-workers are authentically sexist, racist or ageist. What matters is that they behave with civility and tolerance, obey the rules of social interaction and are sincere about it. The criteria of sincerity are unambiguous: Will they keep their promises? Will they honor the meanings and understandings we tacitly negotiate? Are their gestures of cordiality offered in conscious good faith?...Sincerity rests in reconciling our performance of tolerance with the people we become. And what it means for us today is that the best way of living in our diverse and contentiously free society is neither to obsess about the hidden depths of our prejudices nor to deny them, but to behave as if we had none."
Patterson suggests that "Authenticity now dominates our way of viewing ourselves and our relationships, with baleful consequences. Within sensitive individuals it breeds doubt; between people it promotes distrust; within groups it enhances group-think in the endless quest to be one with the group’s true soul; and between groups it is the inner source of identity politics...the primacy of the self has penetrated major areas of government: emotivist arguments trump reasoned discourse in Congressional hearings and criminal justice; and in public education."
"Social scientists and pollsters routinely belittle results showing growing tolerance; they argue that Americans have simply learned how to conceal their deeply...Harvard social psychologist Mahzarin Banaji and her collaborators claim to have evidence, based on more than three million self-administered Web-based tests, that nearly all of us are authentically bigoted to the core with hidden “implicit prejudices” — about race, gender, age, homosexuality and appearance — that we deny, sometimes with consciously tolerant views ingrained prejudices."
"I couldn’t care less whether my neighbors and co-workers are authentically sexist, racist or ageist. What matters is that they behave with civility and tolerance, obey the rules of social interaction and are sincere about it. The criteria of sincerity are unambiguous: Will they keep their promises? Will they honor the meanings and understandings we tacitly negotiate? Are their gestures of cordiality offered in conscious good faith?...Sincerity rests in reconciling our performance of tolerance with the people we become. And what it means for us today is that the best way of living in our diverse and contentiously free society is neither to obsess about the hidden depths of our prejudices nor to deny them, but to behave as if we had none."
Tuesday, January 02, 2007
Explaining away the supernatural as brain misfirings?
Several of my posts have mentioned work suggesting a basis for phenomena like out of body experiences or sensing the presence of phantom others: in the temporary perturbation of brain processes that normally arrange our perception of the external world and others in it. These perturbations have been observed during epileptic seizures and electrical or magnetic stimulation of regions of the brain. Deborah Blum, author of “Ghost Hunters: William James and the Scientific Search for Life After Death.” weighs in on this issue in a Op-Ed piece in the Dec. 30 New York Times. She cites work from Blanke's laboratory that I mentioned in my Oct. 3 post. She seems critical of scientists who "concluded that ghosts are mere “bodily delusions,” electrical misfirings and nothing more" and cites work done on psychic phenomena by respected scientists in the late 19th century. Blum says "Dr. Blanke believes that even this one subject’s experience serves as an example of how we may mistake errant signals in the brain for something more. Humans tend, he points out, to seek explanation, to impose meaning on events that may have none. The pure rationalists among us suggest that our need to add meaning to a basic, biological existence easily accounts for the way we organize religions and find evidence of otherworldly powers in the stuff of everyday life."
Blum then continues: "The nonpurists suggest a different conclusion: willful scientific blindness. And there’s no reason Dr. Blanke’s study can’t support their theories of the paranormal. Perhaps his experimental electric current simply mimics the work of an equally powerful spirit. Much of the psychical research done today applies similar principles: brain-imaging machines highlight parts of the brain that respond to psychic phenomena, while other devices are used to search for infrared radiation or increased electrical activity in haunted houses."
Wait a minute... Equally powerful spirit? Will someone please measure this spirit with a physical instrument, because it is altering physical processes in the brain! Or, "parts of the brain that respond to psychic phenomena?" What is cause and what is effect here? Are we presupposing the existence of psychic phenomena as causes? Then please measure them. I'm sorry, but I can't give up my skepticism about things that alter material physical processes in the brain, how can a non-physical process (spirit, ectoplasm, soul, whatever) change them? We're back to Descartes putting the soul in the pineal gland.

Photo credit: New York Times.
Blum then continues: "The nonpurists suggest a different conclusion: willful scientific blindness. And there’s no reason Dr. Blanke’s study can’t support their theories of the paranormal. Perhaps his experimental electric current simply mimics the work of an equally powerful spirit. Much of the psychical research done today applies similar principles: brain-imaging machines highlight parts of the brain that respond to psychic phenomena, while other devices are used to search for infrared radiation or increased electrical activity in haunted houses."
Wait a minute... Equally powerful spirit? Will someone please measure this spirit with a physical instrument, because it is altering physical processes in the brain! Or, "parts of the brain that respond to psychic phenomena?" What is cause and what is effect here? Are we presupposing the existence of psychic phenomena as causes? Then please measure them. I'm sorry, but I can't give up my skepticism about things that alter material physical processes in the brain, how can a non-physical process (spirit, ectoplasm, soul, whatever) change them? We're back to Descartes putting the soul in the pineal gland.
Photo credit: New York Times.
Thursday, November 02, 2006
Self recognition - humans, apes, dolphins...and now elephants
Plotnik, de Waal, and Reiss report in PNAS that mirror self recognition (MSR), considered an indicator of self awareness, can be observed in Asian elephants. Slightly edited from their abstract: "MSR is thought to correlate with higher forms of empathy and altruistic behavior. Apart from humans and apes, dolphins and elephants are also known for such capacities. After the recent discovery of MSR in dolphins (Tursiops truncatus), elephants thus were the next logical candidate species. The authors exposed three Asian elephants (Elephas maximus) to a large mirror to investigate their responses. Animals that possess MSR typically progress through four stages of behavior when facing a mirror: (i) social responses, (ii) physical inspection (e.g., looking behind the mirror), (iii) repetitive mirror-testing behavior, and (iv) realization of seeing themselves. Visible marks and invisible sham-marks were applied to the elephants' heads to test whether they would pass the litmus "mark test" for MSR in which an individual spontaneously uses a mirror to touch an otherwise imperceptible mark on its own body. Here, we report a successful MSR elephant study and report striking parallels in the progression of responses to mirrors among apes, dolphins, and elephants. These parallels suggest convergent cognitive evolution most likely related to complex sociality and cooperation.

Above video (click the rectangle to play) was taken from the Elmo lipstick camera embedded in the mirror on Happy's first day of marking. Happy repetitively touches the mark or the area immediately around the mark with her trunk while in full view of the mirror. She never touches the sham-mark during this session.
Above video (click the rectangle to play) was taken from the Elmo lipstick camera embedded in the mirror on Happy's first day of marking. Happy repetitively touches the mark or the area immediately around the mark with her trunk while in full view of the mirror. She never touches the sham-mark during this session.
Tuesday, October 10, 2006
A striking difference in brain function in autism: Failure to deactivate.
Kennedy et al report interesting functional magnetic resonance imaging (fMRI) data on normal compared with autistic brains. From their article:
Internally directed processes, such as self-reflective thought and most higher-order social and emotional processes, consistently activate a medial cortical network involving several brain regions, namely, the medial prefrontal cortex (MPFC) and adjacent rostral anterior cingulate cortex (rACC), posterior cingulate cortex (PCC), and precuneus (PrC). Interestingly, this network is active when normal subjects are passively resting, leading many to speculate that these internally directed thoughts dominate the resting state. Self-reports from subjects while at rest further support this interpretation, wherein they typically describe "autobiographical reminiscences, either recent or ancient, consisting of familiar faces, scenes, dialogues, stories, and melodies". Conversely, activity in this midline "resting network" is reduced when subjects perform externally directed, attention-demanding, goal-oriented tasks (such as the Stroop task or math calculations), and the resulting "deactivation" of this network is thought to be an indicator of an interruption of ongoing internally directed thought processes. Thus, measuring deactivation provides a means by which rest-associated functional activity can be quantitatively examined.
Applying this approach to autism, Kennedy et al found that the autism group failed to demonstrate this deactivation effect. Furthermore, there was a strong correlation between a clinical measure of social impairment and functional activity within the ventral medial prefrontal cortex. They speculate that the lack of deactivation in the autism group is indicative of abnormal internally directed processes at rest, which may be an important contribution to the social and emotional deficits of autism.
Internally directed processes, such as self-reflective thought and most higher-order social and emotional processes, consistently activate a medial cortical network involving several brain regions, namely, the medial prefrontal cortex (MPFC) and adjacent rostral anterior cingulate cortex (rACC), posterior cingulate cortex (PCC), and precuneus (PrC). Interestingly, this network is active when normal subjects are passively resting, leading many to speculate that these internally directed thoughts dominate the resting state. Self-reports from subjects while at rest further support this interpretation, wherein they typically describe "autobiographical reminiscences, either recent or ancient, consisting of familiar faces, scenes, dialogues, stories, and melodies". Conversely, activity in this midline "resting network" is reduced when subjects perform externally directed, attention-demanding, goal-oriented tasks (such as the Stroop task or math calculations), and the resulting "deactivation" of this network is thought to be an indicator of an interruption of ongoing internally directed thought processes. Thus, measuring deactivation provides a means by which rest-associated functional activity can be quantitatively examined.
Applying this approach to autism, Kennedy et al found that the autism group failed to demonstrate this deactivation effect. Furthermore, there was a strong correlation between a clinical measure of social impairment and functional activity within the ventral medial prefrontal cortex. They speculate that the lack of deactivation in the autism group is indicative of abnormal internally directed processes at rest, which may be an important contribution to the social and emotional deficits of autism.
Wednesday, October 04, 2006
Clever experiment: distinction of self and other in mirroring motor neurons.
Extravagant claims have been made about systems of neurons that are active both during execution of a motion or emotion and observing others doing the same thing. (Example: Ramachandran's "Mirrors Neurons will do for psychology what DNA did for Biology."). They are suggested to be a basis of empathy and the development of language.
The fact that the brain might represent others' actions like one's own raise the issue of how we distinguish self from other. What keeps us from constantly miming the actions of others? (This happens in echopraxia, the involuntary repetition or imitation of the observed movements of others.) Schütz-Bosbach et al have done a very clever experiment to examine this by manipulating the sense of body ownership (using the “rubber-hand illusion”) to compare effects of observing actions that either were or were not illusorily attributed to the subject's own body.

"When subjects watch a rubber hand being stroked while they feel synchronous stroking of their own unseen hand, they feel that the rubber hand becomes part of their body. Identical asynchronous stroking has no effect. Thus, the sense of owning the rubber hand requires congruence of visual and tactile stimulation. The neural counterparts of this sense of ownership have been identified in premotor and sensorimotor cortices. The rubber-hand illusion therefore allows balanced comparison between the self and the other because a single stimulus (here, the hand of another person rather than a rubber hand) is either linked to the self or not depending on the pattern of previous stimulation. We used a real human hand instead of the conventional rubber hand because several studies show stronger mirroring effects for viewing a live action than for viewing artificial equivalents."
They show that observing another's actions facilitated the motor system, whereas observing identical actions, which were illusorily attributed to the subject's own body, showed the opposite pattern. Thus, motor facilitation strongly depends on the agent to whom the observed action is attributed. This result contradicts previous concepts of equivalence between one's own actions and actions of others and suggests that social differentiation, not equivalence, is characteristic of the human action system.... "This suggests that the neural mechanisms underlying action observation are intrinsically social. These mechanisms map the actions of others to corresponding actions on one's own body but do not simply represent the other agent as a derivative of, or even an equal to, the self." In contrast, there appears to be an agent-specific representation in the primary motor cortex.
The fact that the brain might represent others' actions like one's own raise the issue of how we distinguish self from other. What keeps us from constantly miming the actions of others? (This happens in echopraxia, the involuntary repetition or imitation of the observed movements of others.) Schütz-Bosbach et al have done a very clever experiment to examine this by manipulating the sense of body ownership (using the “rubber-hand illusion”) to compare effects of observing actions that either were or were not illusorily attributed to the subject's own body.

"When subjects watch a rubber hand being stroked while they feel synchronous stroking of their own unseen hand, they feel that the rubber hand becomes part of their body. Identical asynchronous stroking has no effect. Thus, the sense of owning the rubber hand requires congruence of visual and tactile stimulation. The neural counterparts of this sense of ownership have been identified in premotor and sensorimotor cortices. The rubber-hand illusion therefore allows balanced comparison between the self and the other because a single stimulus (here, the hand of another person rather than a rubber hand) is either linked to the self or not depending on the pattern of previous stimulation. We used a real human hand instead of the conventional rubber hand because several studies show stronger mirroring effects for viewing a live action than for viewing artificial equivalents."
They show that observing another's actions facilitated the motor system, whereas observing identical actions, which were illusorily attributed to the subject's own body, showed the opposite pattern. Thus, motor facilitation strongly depends on the agent to whom the observed action is attributed. This result contradicts previous concepts of equivalence between one's own actions and actions of others and suggests that social differentiation, not equivalence, is characteristic of the human action system.... "This suggests that the neural mechanisms underlying action observation are intrinsically social. These mechanisms map the actions of others to corresponding actions on one's own body but do not simply represent the other agent as a derivative of, or even an equal to, the self." In contrast, there appears to be an agent-specific representation in the primary motor cortex.
Tuesday, October 03, 2006
Your brain can put your body wherever it likes....
Increasingly there seems to be little point in hanging on to new age spiritual, paranormal, or esoteric fantasies about altered mental states in which we seem to leave our bodies. We're actually not going anywhere... our brains are just making it up. We can, readily alter our sense of body ownership, as in the well known "rubber hand" illusion.
A nice graphic in today's New York Times prompts me to go ahead and mention a recent report from Blanke and coworkers in Nature that I was going to pass by, because it has received wide notice in the press and other blogs. This paper adds to a growing literature whose bottom line is that perturbation of nerve firing in brain areas near where the temporal and parietal lobes meet can cause a variety of distortions of our subjective sense of our body in space. Most commonly this is felt as an "out of body experience" where we are looking at ourselves from some external perspective or experiencing a 'shadow' version of ourselves. Here is the graphic from the New York Times based on work from the Blanke laboratory:

(Click to enlarge)
A nice graphic in today's New York Times prompts me to go ahead and mention a recent report from Blanke and coworkers in Nature that I was going to pass by, because it has received wide notice in the press and other blogs. This paper adds to a growing literature whose bottom line is that perturbation of nerve firing in brain areas near where the temporal and parietal lobes meet can cause a variety of distortions of our subjective sense of our body in space. Most commonly this is felt as an "out of body experience" where we are looking at ourselves from some external perspective or experiencing a 'shadow' version of ourselves. Here is the graphic from the New York Times based on work from the Blanke laboratory:

(Click to enlarge)
Wednesday, September 27, 2006
Brain correlates of hysteria.
The Tuesday Science section of the New York Times (Sept 26) has an interesting article on hysteria, a fashionable syndrome in the Victorian era which has "disappeared" during this century. Actually the term "conversion disorder" is now used to describe an ill-defined syndrome with no obvious physical cause, usually involving paralysis of a portion of the body or seizures. Sigmund Freud suggested from his case studies that hysteria is something in the psyche or the mind being expressed physically in the body.

The 19th-century French neurologist Jean-Martin Charcot, shown lecturing on hysteria
Peter W. Halligan at Cardiff, co-founder of the journal Cognitive Neuropsychiatry and his colleagues "analyzed the brain function of a woman who was paralyzed on the left side of her body (Cognition, 64, B1-B8, 1997). First they conducted numerous tests to ensure that she had no identifiable organic lesion...When the woman tried to move her “paralyzed leg,” her primary motor cortex was not activated as it should have been; instead her right orbitofrontal and right anterior cingulate cortex, parts of the brain that have been associated with action and emotion, were activated. They reasoned that these emotional areas of the brain were responsible for suppressing movement in her paralyzed leg."


Fig. 1. Relative rCBF (blood flow measured by magnetic resonance imaging) increases associated with movement of the right (good) leg. The figure reveals relative rCBF increases when the normal (right) leg is moved that do not occur when attempts to move the bad (left) leg are made. There is left hemisphere neuronal activation centered on the primary sensory and motor cortex. Additional activation is seen in the left inferior parietal cortex and the right inferior temporal cortex.
Fig. 2. Relative rCBF increases associated with attempted movement of the left (bad) leg. This reveals relative rCBF increases during attempts to move the bad (left) leg that did not occur when the good (right) leg was moved. There is activation in the right anterior cingulate and the right orbito-frontal cortex.
“The patient willed her leg to move,” Dr. Halligan said. “But that act of willing triggered this primitive orbitofrontal area and activated the anterior cingulate to countermand the instruction to move the leg. She was willing it, but the leg would not move.”
"Subsequent studies have bolstered the notion that parts of the brain involved in emotion may be activated inappropriately in patients with conversion disorder and may inhibit the normal functioning of brain circuitry responsible for movement, sensation and sight......Both its persistence and its pervasiveness suggest that hysteria may be derived from an instinctual response to threat. Total shutdown, in the form of paralysis, for example, is not an entirely untoward or unheard of response to an untenable situation. (Think of deer in the headlights.)"

The 19th-century French neurologist Jean-Martin Charcot, shown lecturing on hysteria
Peter W. Halligan at Cardiff, co-founder of the journal Cognitive Neuropsychiatry and his colleagues "analyzed the brain function of a woman who was paralyzed on the left side of her body (Cognition, 64, B1-B8, 1997). First they conducted numerous tests to ensure that she had no identifiable organic lesion...When the woman tried to move her “paralyzed leg,” her primary motor cortex was not activated as it should have been; instead her right orbitofrontal and right anterior cingulate cortex, parts of the brain that have been associated with action and emotion, were activated. They reasoned that these emotional areas of the brain were responsible for suppressing movement in her paralyzed leg."


Fig. 1. Relative rCBF (blood flow measured by magnetic resonance imaging) increases associated with movement of the right (good) leg. The figure reveals relative rCBF increases when the normal (right) leg is moved that do not occur when attempts to move the bad (left) leg are made. There is left hemisphere neuronal activation centered on the primary sensory and motor cortex. Additional activation is seen in the left inferior parietal cortex and the right inferior temporal cortex.
Fig. 2. Relative rCBF increases associated with attempted movement of the left (bad) leg. This reveals relative rCBF increases during attempts to move the bad (left) leg that did not occur when the good (right) leg was moved. There is activation in the right anterior cingulate and the right orbito-frontal cortex.
“The patient willed her leg to move,” Dr. Halligan said. “But that act of willing triggered this primitive orbitofrontal area and activated the anterior cingulate to countermand the instruction to move the leg. She was willing it, but the leg would not move.”
"Subsequent studies have bolstered the notion that parts of the brain involved in emotion may be activated inappropriately in patients with conversion disorder and may inhibit the normal functioning of brain circuitry responsible for movement, sensation and sight......Both its persistence and its pervasiveness suggest that hysteria may be derived from an instinctual response to threat. Total shutdown, in the form of paralysis, for example, is not an entirely untoward or unheard of response to an untenable situation. (Think of deer in the headlights.)"
Tuesday, September 12, 2006
God on the Brain

The Economist has a nice review and critical discussion of work by Mario Beauregard and collaborators at the Univ. of Montreal doing fMRI imaging of the brains of Carmelite nuns as they recall experiences of mystical union (by definition such experiences can not be summoned at will). The idea is based on the fact that imagining an experience usually activates the same brain regions that are active when the experience is actually taking place. Not surprisingly, there is no "God spot" in the brain, and activity in a number of brain regions, notably emotional areas, correlates with the recall of union. From their abstract in Neuroscience Letters (Volume 405, Issue 3 , 25 September 2006): "The brain activity of Carmelite nuns was measured while they were subjectively in a state of union with God. This state was associated with significant loci of activation in the right medial orbitofrontal cortex, right middle temporal cortex, right inferior and superior parietal lobules, right caudate, left medial prefrontal cortex, left anterior cingulate cortex, left inferior parietal lobule, left insula, left caudate, and left brainstem. Other loci of activation were seen in the extra-striate visual cortex. These results suggest that mystical experiences are mediated by several brain regions and systems."
Brain imaging has been to used to study a number of other altered states of consciousness, such as the phantom limb phenomenon and out of body experiences. Brain damage in the region of junction of the temporal and parietal lobes can alter perception of personal and extrapersonal space, and other studies have shown the changes in activation in this region correlates with meditative experiences of sensing sensing a greater interconnectedness of things, and dissolution of self into some larger entity.
Wednesday, September 06, 2006
Neural operations that give rise to a unitary sense of self.
An interesting article by Moran et. al. in Journal of Cognitive Neuroscience: They examined whether the cognitive and affective components of self-reflection can be dissociated using functional magnetic resonance imaging. Using a simple paradigm in which subjects judged the personal relevance of personality characteristics that were either favorable (e.g., "honest") or unfavorable (e.g., "lazy"), they found that distinct neural circuits in adjacent regions of the prefrontal cortex subserve cognitive and emotional aspects of self-reflection. The medial prefrontal cortex responded only to material that was self-descriptive, and this did not differ as a function of the valence of the trait. When material was judged to be self-relevant, the valence of the material was resolved in an adjacent region of ventral anterior cingulate.

Figure Legend: Whole-brain ANOVA analysis revealed a main effect of self-relevance (top left) in the medial prefrontal cortex (MPFC) and posterior cingulate cortex (pCC), a main effect of valence (top right) in the ventral anterior cingulate cortex (vACC), and a self-relevance by valence interaction (bottom left) in the vACC, the dorsal anterior cingulate cortex (dACC), and the supplementary motor area (SMA). To qualitatively identify whether brain regions identified in the ANOVA analysis showed a strong bias toward one of the two main effects, a self-relevance/valence sensitivity measure (F ratio) was computed on a voxel-by-voxel basis by dividing the self-relevance F score for each voxel by the valence F score. Voxels that did not yield a significant main effect of either self-relevance or valence were excluded from further analysis to avoid spurious F ratio effects. To facilitate visualization of this sensitivity measure, F ratios were transformed to a logarithmic scale. Voxels that were more sensitive to trait valence yielded negative values (blue color scale), whereas voxels that were more sensitive to self-relevance yielded positive values (yellow color scale). Voxels at the tail end of the color scales were those voxels that exhibited the greatest bias toward trait valence and self-relevance, respectively. Voxels in the MPFC (BA 10) and two regions of the pCC (BA 29/30 and BA 23) demonstrated greater sensitivity to self-relevance, whereas voxels in the vACC (BA 25) demonstrated greater sensitivity to trait valence.

Figure Legend: Whole-brain ANOVA analysis revealed a main effect of self-relevance (top left) in the medial prefrontal cortex (MPFC) and posterior cingulate cortex (pCC), a main effect of valence (top right) in the ventral anterior cingulate cortex (vACC), and a self-relevance by valence interaction (bottom left) in the vACC, the dorsal anterior cingulate cortex (dACC), and the supplementary motor area (SMA). To qualitatively identify whether brain regions identified in the ANOVA analysis showed a strong bias toward one of the two main effects, a self-relevance/valence sensitivity measure (F ratio) was computed on a voxel-by-voxel basis by dividing the self-relevance F score for each voxel by the valence F score. Voxels that did not yield a significant main effect of either self-relevance or valence were excluded from further analysis to avoid spurious F ratio effects. To facilitate visualization of this sensitivity measure, F ratios were transformed to a logarithmic scale. Voxels that were more sensitive to trait valence yielded negative values (blue color scale), whereas voxels that were more sensitive to self-relevance yielded positive values (yellow color scale). Voxels at the tail end of the color scales were those voxels that exhibited the greatest bias toward trait valence and self-relevance, respectively. Voxels in the MPFC (BA 10) and two regions of the pCC (BA 29/30 and BA 23) demonstrated greater sensitivity to self-relevance, whereas voxels in the vACC (BA 25) demonstrated greater sensitivity to trait valence.
Wednesday, August 16, 2006
The Neural Basis of Embodyment
Some edited clips from a recent J. Neuroscience article by Arzy et al. :
Embodiment, the sense of being localized within one's physical body, is a fundamental aspect of the self. Recent evidence has started to show that self and body processing require two distinct brain mechanisms, with key loci in the temporoparietal junction (TPJ) - involved in self processing and multisensory integration of body-related information - and the extrastriate body area (EBA) - which responds selectively to human bodies and body parts. Arzy et al have used evoked potential mapping to show that activations in EBA and TPJ code differentially for embodiment and self location, because the location and timing of brain activation depended on whether mental imagery is performed with mentally embodied (EBA) or disembodied (TPJ) self location. In a second experiment, they showed that only EBA activation, related to embodied self location, but not TPJ activation, related to disembodied self location, was modified by the subjects' body position during task performance (supine or sitting). This suggests that embodied self location and actual body location share neural mechanisms.

Figure. To investigate embodiment and self location, subjects were asked to perform two mental-imagery tasks with respect to their own body in response to a schematic front- or back-facing human figure. In an own-body transformation task, (OBT task) subjects were asked to imagine themselves in the position and orientation of a schematic human figure, as shown on a computer screen (bottom row, the correct responses for each task are indicated under each stimulus) Either the right or left hand of the figure was marked, and subjects indicated which hand was marked. In a mirror task (MIR task), the same schematic human figure was shown, but subjects were instructed to imagine that the schematic figure (as shown on the computer screen) was their mirror reflection, as seen from their habitual point of view ( top row, the correct responses for each task are indicated under each stimulus).

Figure. Generators of mirror task (MIR. top row, were localized at the left EBA and of the own body transformation task (OBT, bottom row) at the right TPJ and left EBA

Figure. EBA. The mean (x, y) Talairach coordinates of the EBA are given for several neuroimaging studies. Note the similarity of EBA localization across studies, neuroimaging techniques, and behavioral tasks.
Embodiment, the sense of being localized within one's physical body, is a fundamental aspect of the self. Recent evidence has started to show that self and body processing require two distinct brain mechanisms, with key loci in the temporoparietal junction (TPJ) - involved in self processing and multisensory integration of body-related information - and the extrastriate body area (EBA) - which responds selectively to human bodies and body parts. Arzy et al have used evoked potential mapping to show that activations in EBA and TPJ code differentially for embodiment and self location, because the location and timing of brain activation depended on whether mental imagery is performed with mentally embodied (EBA) or disembodied (TPJ) self location. In a second experiment, they showed that only EBA activation, related to embodied self location, but not TPJ activation, related to disembodied self location, was modified by the subjects' body position during task performance (supine or sitting). This suggests that embodied self location and actual body location share neural mechanisms.

Figure. To investigate embodiment and self location, subjects were asked to perform two mental-imagery tasks with respect to their own body in response to a schematic front- or back-facing human figure. In an own-body transformation task, (OBT task) subjects were asked to imagine themselves in the position and orientation of a schematic human figure, as shown on a computer screen (bottom row, the correct responses for each task are indicated under each stimulus) Either the right or left hand of the figure was marked, and subjects indicated which hand was marked. In a mirror task (MIR task), the same schematic human figure was shown, but subjects were instructed to imagine that the schematic figure (as shown on the computer screen) was their mirror reflection, as seen from their habitual point of view ( top row, the correct responses for each task are indicated under each stimulus).

Figure. Generators of mirror task (MIR. top row, were localized at the left EBA and of the own body transformation task (OBT, bottom row) at the right TPJ and left EBA

Figure. EBA. The mean (x, y) Talairach coordinates of the EBA are given for several neuroimaging studies. Note the similarity of EBA localization across studies, neuroimaging techniques, and behavioral tasks.
Friday, August 11, 2006
"Hare Brain, Tortoise Mind" and "The Wayward Mind"
These are the titles of two books by British psychologist Guy Claxton (see his website) that I think have received less attention than they should. (It seems to me that British and American psychologists group themselves in quite separate worlds.) The first book is a lucid presentation of experimental work that supports Herbert Spenser's dictum "The determined effort causes perversion of thought." Claxton uses the term "undermind" to describe intuitive and integrative processes normally beyond the range of, and can be inhibited by, our focused awareness. Extremes of being indiscriminately intuitive or insisting on lots of high-quality information can block results.
The term 'undermind' hasn't caught on, and the subsequent excellent book "The Wayward Mind" reverts to using the term "unconscious". "The Wayward Mind" is a history of human attempts to explain the unconscious mind, from ancient descriptions of the 'underworld' to the theories of modern neuroscience.
Here are (clipped and truncated) some lines from pp 348-352 of "Wayward Mind" that I like:
"What we call our ‘self’ is an agglomeration of both conscious and unconscious ingredients: cans, needs, dos, oughts, thinks….these constructions hold out an overwhelming temptation: to assume that the “I” is the same in all of them… so that instead of having an intricate web of things that make me Me, I have to create a single imaginary hub around which they all revolve, to which they all refer…the attempt to keep this fiction going, to ‘hold it together’ can become quite tiring and bothersome… If “I” am essentially reasonable, if I imagine that my zones of control – over my own feelings for example – are wider and more robust than they are, then I am going to get in a tangle trying to ‘control myself.’ If I have decided that who I am is clever, attractive, athletic, stable, creating the hub of “I” locks everything together and prevents it moving. It stops Me expanding to include the unconscious, or graciously shrinking to accommodate old age. I can’t enjoy my waywardness, nor see it as an intrinsic part of ME….All the evidence is that a more relaxed attitude toward the bounds of self makes for a richer, easier and more creative life. Perhaps, after all, waywardness in all its forms is in need not so much of explanation, but of a mystified but friendly welcome. We can explain it if we wish, and the brain is beginning to a reasonable job. But the need to explain, when not motivated by the dispassionate curiosity of the scientist, is surely a sign of anxiety: of the desire to tame with words that which is experienced as unsettling.. "
The term 'undermind' hasn't caught on, and the subsequent excellent book "The Wayward Mind" reverts to using the term "unconscious". "The Wayward Mind" is a history of human attempts to explain the unconscious mind, from ancient descriptions of the 'underworld' to the theories of modern neuroscience.
Here are (clipped and truncated) some lines from pp 348-352 of "Wayward Mind" that I like:
"What we call our ‘self’ is an agglomeration of both conscious and unconscious ingredients: cans, needs, dos, oughts, thinks….these constructions hold out an overwhelming temptation: to assume that the “I” is the same in all of them… so that instead of having an intricate web of things that make me Me, I have to create a single imaginary hub around which they all revolve, to which they all refer…the attempt to keep this fiction going, to ‘hold it together’ can become quite tiring and bothersome… If “I” am essentially reasonable, if I imagine that my zones of control – over my own feelings for example – are wider and more robust than they are, then I am going to get in a tangle trying to ‘control myself.’ If I have decided that who I am is clever, attractive, athletic, stable, creating the hub of “I” locks everything together and prevents it moving. It stops Me expanding to include the unconscious, or graciously shrinking to accommodate old age. I can’t enjoy my waywardness, nor see it as an intrinsic part of ME….All the evidence is that a more relaxed attitude toward the bounds of self makes for a richer, easier and more creative life. Perhaps, after all, waywardness in all its forms is in need not so much of explanation, but of a mystified but friendly welcome. We can explain it if we wish, and the brain is beginning to a reasonable job. But the need to explain, when not motivated by the dispassionate curiosity of the scientist, is surely a sign of anxiety: of the desire to tame with words that which is experienced as unsettling.. "
Thursday, August 03, 2006
Personality Predicts Responsivity of the Brain Reward System
The Journal of Neuroscience has started a new feature: short reviews of a recent paper in the Journal, written exclusively by graduate students or postdoctoral fellows. They are meant to mimic the journal clubs that exist in many academic department or institutions.
A recent entry by J. B. Engelmann discusses a paper by Beaver et al. that examines the relationship between a personality trait, reward sensitivity, and activity of the brain reward system measured by functional magnetic resonance imaging. A study like this is relevant to understanding the over-consumption of appetizing high-caloric foods that has contributed to the dramatic increase in obesity within the past 20 years, making obesity a top 10 global health threat. Here are some clips from the review and article:

Figure - A highly simplified schematic diagram outlines the connections between central nodes in the brain reward system (modified from Berridge and Robinson, 2003). Animal as well as human neuroimaging studies have implicated this network in the hedonic and motivational effects of natural rewards and drugs of abuse, such as food and amphetamine. AMYG, Amygdala; NAC, nucleus accumbens; OFC, orbitofrontal cortex; VP, ventral pallidum; VTA/SN, ventral tegmental area/substantia nigra.
Beaver et al. showed that reward sensitivity, as assessed by the Behavioral Inhibition Scale/Behavioral Activation Scale, predicted neural responses to pictures of appetizing foods in the network of the brain regions outlined in the Figure. Functional magnetic resonance imaging, in conjunction with a blocked experimental design, was used to record blood oxygenation-level dependent (BOLD) responses while participants passively viewed pictures of foods from four different categories...The authors found increased activation in orbitofrontal cortex (appetizing vs nonfood objects) and bilateral ventral striatum (appetizing vs bland foods). Interestingly, there was a dissociation between left and right orbitofrontal cortex such that appetizing food stimuli activated the left orbitofrontal cortex, whereas disgusting food stimuli activated the right orbitofrontal cortex.
Beaver et al. provide an important link between human behavioral research that has demonstrated an association between trait reward sensitivity and unhealthy eating habits and animal research implicating the reward network in hyperphagia and increased intake of high-caloric foods. Their findings thus offer a possible explanation for why some individuals overeat compulsively. It will be interesting to establish a more direct link between compulsive overeating and responsivity of the reward system, for instance by correlating activity in the reward system in response to images depicting appetizing foods and body mass index. Finally, an abundance of parallels between obesity and drug addiction points to similarities in the underlying brain mechanisms and neural adaptations that accompany these two conditions
A recent entry by J. B. Engelmann discusses a paper by Beaver et al. that examines the relationship between a personality trait, reward sensitivity, and activity of the brain reward system measured by functional magnetic resonance imaging. A study like this is relevant to understanding the over-consumption of appetizing high-caloric foods that has contributed to the dramatic increase in obesity within the past 20 years, making obesity a top 10 global health threat. Here are some clips from the review and article:

Figure - A highly simplified schematic diagram outlines the connections between central nodes in the brain reward system (modified from Berridge and Robinson, 2003). Animal as well as human neuroimaging studies have implicated this network in the hedonic and motivational effects of natural rewards and drugs of abuse, such as food and amphetamine. AMYG, Amygdala; NAC, nucleus accumbens; OFC, orbitofrontal cortex; VP, ventral pallidum; VTA/SN, ventral tegmental area/substantia nigra.
Beaver et al. showed that reward sensitivity, as assessed by the Behavioral Inhibition Scale/Behavioral Activation Scale, predicted neural responses to pictures of appetizing foods in the network of the brain regions outlined in the Figure. Functional magnetic resonance imaging, in conjunction with a blocked experimental design, was used to record blood oxygenation-level dependent (BOLD) responses while participants passively viewed pictures of foods from four different categories...The authors found increased activation in orbitofrontal cortex (appetizing vs nonfood objects) and bilateral ventral striatum (appetizing vs bland foods). Interestingly, there was a dissociation between left and right orbitofrontal cortex such that appetizing food stimuli activated the left orbitofrontal cortex, whereas disgusting food stimuli activated the right orbitofrontal cortex.
Beaver et al. provide an important link between human behavioral research that has demonstrated an association between trait reward sensitivity and unhealthy eating habits and animal research implicating the reward network in hyperphagia and increased intake of high-caloric foods. Their findings thus offer a possible explanation for why some individuals overeat compulsively. It will be interesting to establish a more direct link between compulsive overeating and responsivity of the reward system, for instance by correlating activity in the reward system in response to images depicting appetizing foods and body mass index. Finally, an abundance of parallels between obesity and drug addiction points to similarities in the underlying brain mechanisms and neural adaptations that accompany these two conditions
Friday, July 21, 2006
A ‘Senior Moment’ or a Self-Fulfilling Prophecy?
This is the title of an article by Benedict Carey in last Tuesday's NYTimes science section (7/18/06) pointing to work reported in the current issue of the journal Social Cognition. Men and women in late middle age (48-62) underperformed on a standard memory test when told they were part of a study including people over age 70. Inclusion with an older group — an indirect reminder of the link between age and memory slippage — was enough to affect their scores, especially for those who were most concerned about getting older. Scores were higher when participants were told they were competing with a group in their 20's. The findings “show how negative images of aging on TV, in other media and in jokes reinforce negative stereotypes that can affect performance even before people reach retirement age."
This self-undermining is a stereotype effect, of the sort that has been documented it in many groups. Other studies have shown that women perform less well on math exams after reading that men tend to perform better on them. Similarly, white men perform less well when they are told that they are competing in math against Asian students.
This self-undermining is a stereotype effect, of the sort that has been documented it in many groups. Other studies have shown that women perform less well on math exams after reading that men tend to perform better on them. Similarly, white men perform less well when they are told that they are competing in math against Asian students.
Tuesday, July 18, 2006
Free Will, Free Won't, or Neither? A refinement of Libet's work on the conscious control of spontaneous actions.
In a famous paper published in 1983, Libet et al. showed that the recordable cerebral activity (readiness-potential) that precedes a freely voluntary motor act occurs at least several hundred milliseconds before the reported time of conscious intention to act. The actual movement occurs 200-250 msec after the reported time of intention to act. The data are pretty spooky when you think about it. They say that your brain ("it") has started working on a action well before "you" think you are initiating it! This article has sparked a continuing debate on whether we actually have free will. Libet has suggested that the ~200-250 msec period between awareness of intention and the actual action was sufficient to permit a "veto" of the action if it was judged inappropriate. In this interpretation, we might be said to be "free won't" rather than "free will".
Lau et al. have now done a more nuanced version of LIbet's experiments. In a previous paper they showed that, when participants were required to estimate the onset of their intentions using Libet's procedure, the activity in the presupplementary motor area (pre-SMA) was enhanced ~228 msec before motor execution. In their most recent work they show that when participants were required to estimate the onset of their motor executions (instead of their intentions), the activity in the cingulate motor area was enhanced. This latter condition, judged to be more natural and have less task-demanding instructions. The perceived onset of intention could be as late as ~120 msec before the motor execution . "Together, these findings raise the question of whether the conscious control of spontaneous action can be done within a much shorter time window than we had expected, or whether, as suggested by Wegner (The illusion of conscious will Cambridge, MA: MIT Press, 2002), our impression of conscious control is simply illusory."
I think Wegner has it right. His book, and his interpretation of our sense of agency as an after the fact ' emotion of authorship' is a must-read for anyone interested in issues of conscious volition.
Lau et al. have now done a more nuanced version of LIbet's experiments. In a previous paper they showed that, when participants were required to estimate the onset of their intentions using Libet's procedure, the activity in the presupplementary motor area (pre-SMA) was enhanced ~228 msec before motor execution. In their most recent work they show that when participants were required to estimate the onset of their motor executions (instead of their intentions), the activity in the cingulate motor area was enhanced. This latter condition, judged to be more natural and have less task-demanding instructions. The perceived onset of intention could be as late as ~120 msec before the motor execution . "Together, these findings raise the question of whether the conscious control of spontaneous action can be done within a much shorter time window than we had expected, or whether, as suggested by Wegner (The illusion of conscious will Cambridge, MA: MIT Press, 2002), our impression of conscious control is simply illusory."
I think Wegner has it right. His book, and his interpretation of our sense of agency as an after the fact ' emotion of authorship' is a must-read for anyone interested in issues of conscious volition.
Thursday, June 15, 2006
Emotional Communication in Humans and other Primates.
This is the title of a review by Parr et al. (Current Opinion in Neurobiology, vol. 15, pp 716-720, Dec. 2005), whose abstract I include here, with a main point of this post being to show pictures of the similar facial muscle groups used by humans and chimpanzees in emotional communication.

 Credit: Current Opinions in Neurobiology.
Credit: Current Opinions in Neurobiology.
"The social brain hypothesis proposes that large neocortex size in Homonoids evolved to cope with the increasing demands of complex group living and greater numbers of interindividual relationships. Group living requires that individuals communicate effectively about environmental and internal events. Recent data have highlighted the complexity of chimpanzee communication, including graded facial expressions and referential vocalizations. Among Hominoids, elaborate facial communication is accompanied by specializations in brain areas controlling facial movement. Finally, the evolution of empathy, or emotional awareness, might have a neural basis in specialized cells in the neocortex, that is, spindle cells that have been associated with self-conscious emotions, and mirror neurons that have recently been shown to activate in response to communicative facial gestures."
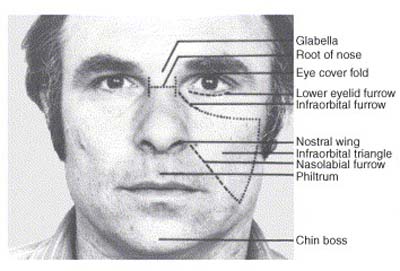
 Credit: Current Opinions in Neurobiology.
Credit: Current Opinions in Neurobiology."The social brain hypothesis proposes that large neocortex size in Homonoids evolved to cope with the increasing demands of complex group living and greater numbers of interindividual relationships. Group living requires that individuals communicate effectively about environmental and internal events. Recent data have highlighted the complexity of chimpanzee communication, including graded facial expressions and referential vocalizations. Among Hominoids, elaborate facial communication is accompanied by specializations in brain areas controlling facial movement. Finally, the evolution of empathy, or emotional awareness, might have a neural basis in specialized cells in the neocortex, that is, spindle cells that have been associated with self-conscious emotions, and mirror neurons that have recently been shown to activate in response to communicative facial gestures."
Thursday, June 08, 2006
Left Brain and Right Brain in the regulation of our subjective feelings.
There is increasing evidence that an the insula areas of our left and right cerebral cortices process higher order re-representations of homeostatic (body regulatory) senses which are sent to prefrontal cortical areas and are the site of our subjective feelings about our bodies. (i.e., sensory information about our bodies from sympathetic and parasympathetic systems feeds to the insula and then on to the prefrontal cortex).
I'm including here quotes from a recent proposal by Craig in Trends in Cognitives Sciences (vol 9,pg. 566,2005) that tries to link many disparate threads to cast an integrated model of how basic regulatory levels of energy expenditure or storage regulated by the sympathetic and parasympathetic nervous systems though out our bodies connect to our higher levels of frontal lobe cognition. His proposal "highlights emerging evidence in support of a direct neuroanatomical relationship in the human forebrain between emotion and homeostasis that mirrors the asymmetric opponent management of energy acquisition and utilization by the autonomic nervous system."
"Recent neurobiological studies using anatomical, neurological, and functional imaging methods indicate that subjectively experienced feelings and emotions might be based on higher-order re-representations of homeostatic afferent (sensory) activity in the human forebrain, and it is particularly noteworthy that such evidence indicates a strong pattern of lateralization. Further, there is a comparable pattern of lateralization evident for the cortical control of cardiac activity, and this can be directly related to left/right asymmetry in the opposing parasympathetic and sympathetic components of the peripheral autonomic nervous system. The confluence of these strikingly parallel asymmetries suggests a homeostatic neuroanatomical model of emotional asymmetry, in which the left forebrain is associated predominantly with parasympathetic activity, and thus with nourishment, safety, positive affect, approach (appetitive) behavior, and group-oriented (affiliative) emotions, while the right forebrain is associated predominantly with sympathetic activity, and thus with arousal, danger, negative affect, withdrawal (aversive) behavior, and individual-oriented (survival) emotions. In the model I am proposing, management of physical and mental (meaning neural) energy is the salient organizational motif, such that energy enrichment is associated with the left forebrain and energy expenditure is associated with the right forebrain, consistent with the respective roles of the parasympathetic and sympathetic efferent systems. The autonomic principle of coordinated opponent interactions between the two hemispheres could provide a fundamental management process."

Figure: Imaging activity in the insula. Credit Univ. of Cambridge Center for Speech and Language.
I'm including here quotes from a recent proposal by Craig in Trends in Cognitives Sciences (vol 9,pg. 566,2005) that tries to link many disparate threads to cast an integrated model of how basic regulatory levels of energy expenditure or storage regulated by the sympathetic and parasympathetic nervous systems though out our bodies connect to our higher levels of frontal lobe cognition. His proposal "highlights emerging evidence in support of a direct neuroanatomical relationship in the human forebrain between emotion and homeostasis that mirrors the asymmetric opponent management of energy acquisition and utilization by the autonomic nervous system."
"Recent neurobiological studies using anatomical, neurological, and functional imaging methods indicate that subjectively experienced feelings and emotions might be based on higher-order re-representations of homeostatic afferent (sensory) activity in the human forebrain, and it is particularly noteworthy that such evidence indicates a strong pattern of lateralization. Further, there is a comparable pattern of lateralization evident for the cortical control of cardiac activity, and this can be directly related to left/right asymmetry in the opposing parasympathetic and sympathetic components of the peripheral autonomic nervous system. The confluence of these strikingly parallel asymmetries suggests a homeostatic neuroanatomical model of emotional asymmetry, in which the left forebrain is associated predominantly with parasympathetic activity, and thus with nourishment, safety, positive affect, approach (appetitive) behavior, and group-oriented (affiliative) emotions, while the right forebrain is associated predominantly with sympathetic activity, and thus with arousal, danger, negative affect, withdrawal (aversive) behavior, and individual-oriented (survival) emotions. In the model I am proposing, management of physical and mental (meaning neural) energy is the salient organizational motif, such that energy enrichment is associated with the left forebrain and energy expenditure is associated with the right forebrain, consistent with the respective roles of the parasympathetic and sympathetic efferent systems. The autonomic principle of coordinated opponent interactions between the two hemispheres could provide a fundamental management process."

Figure: Imaging activity in the insula. Credit Univ. of Cambridge Center for Speech and Language.
Friday, May 19, 2006
Two ways of knowing the minds of others
Mitchell's laboratory at Harvard has shed light on two traditionally opposed hypotheses about how we infer the mental states of others. Simulation theory posits that we use our own experience to infer the experience of others. It is known that when we observe actions and emotions in others, regions in our own brain that would generate those actions or emotions become active and mirror what we are observing. Theory of mind, on the other hand, holds that we use abstract rules about how people behave to infer the mental states of others.
Mitchell et al used functional neuroimaging to examine how perceivers make mental state inferences when such self-other overlap can be assumed (when the other is similar to oneself) and when it cannot (when the other is dissimilar from oneself). "We observed a double dissociation such that mentalizing about a similar other engaged a region of ventral mPFC (medial prefrontal cortex) linked to self-referential thought, whereas mentalizing about a dissimilar other engaged a more dorsal subregion of mPFC. "
 Legend: Division of labor. Different regions of prefrontal cortex fire up when people ponder the mental states of others perceived as similar (blue) or dissimilar (red) to themselves. Credit: Jason Mitchell.
Legend: Division of labor. Different regions of prefrontal cortex fire up when people ponder the mental states of others perceived as similar (blue) or dissimilar (red) to themselves. Credit: Jason Mitchell.
"The overlap between judgments of self and similar others suggests the plausibility of "simulation" accounts of social cognition, which posit that perceivers can use knowledge about themselves to infer the mental states of others." And, the activation of dorsal mPFC during thinking about dissimilar others might correspond to more rule bound theory of mind operations.
Mitchell et al used functional neuroimaging to examine how perceivers make mental state inferences when such self-other overlap can be assumed (when the other is similar to oneself) and when it cannot (when the other is dissimilar from oneself). "We observed a double dissociation such that mentalizing about a similar other engaged a region of ventral mPFC (medial prefrontal cortex) linked to self-referential thought, whereas mentalizing about a dissimilar other engaged a more dorsal subregion of mPFC. "
 Legend: Division of labor. Different regions of prefrontal cortex fire up when people ponder the mental states of others perceived as similar (blue) or dissimilar (red) to themselves. Credit: Jason Mitchell.
Legend: Division of labor. Different regions of prefrontal cortex fire up when people ponder the mental states of others perceived as similar (blue) or dissimilar (red) to themselves. Credit: Jason Mitchell."The overlap between judgments of self and similar others suggests the plausibility of "simulation" accounts of social cognition, which posit that perceivers can use knowledge about themselves to infer the mental states of others." And, the activation of dorsal mPFC during thinking about dissimilar others might correspond to more rule bound theory of mind operations.
Sunday, April 09, 2006
Our inability to cope with what empirical data show us to be true about how our minds work...
I get frustrated when I try to reconcile what I know from empirical data to be true about my self (see the "I-Illusion" essay on this website) with the common sense feeling of agency and responsibility that we are share.
Our commonsense conceptions of ourselves have co-evolved over hundreds of thousands of years, along with their physiological, homeostatic, neuroendocrine, and limbic emotional correlates. This whole complex (us, that is) can be upset by facing what it can come to know to be true about the impersonal physical processes that actually run our show, finding it impossible to integrate its 'illusory' self image.
Here is a clip, and then its more extended context from the piece by Metzinger on edge.org..his response to the question "What is your dangerous idea." He frames it much better than I can. First the clip:
"I think that the irritation and deep sense of resentment surrounding public debates on the freedom of the will actually has nothing much to do with the actual options on the table. It has to do with the perfectly sensible intuition that our presently obvious answer will not only be emotionally disturbing, but ultimately impossible to integrate into our conscious self-models."
Then the more extended quotation:
"For middle-sized objects at 37° like the human brain and the human body, determinism is obviously true. The next state of the physical universe is always determined by the previous state. And given a certain brain-state plus an environment you could never have acted otherwise. A surprisingly large majority of experts in the free-will debate today accept this obvious fact...."
"Yes, you are a physically determined system. But this is not a big problem, because, under certain conditions, we may still continue to say that you are "free": all that matters is that your actions are caused by the right kinds of brain processes and that they originate in you. A physically determined system can well be sensitive to reasons and to rational arguments, to moral considerations, to questions of value and ethics, as long as all of this is appropriately wired into its brain. You can be rational, and you can be moral, as long as your brain is physically determined in the right way. You like this basic idea: physical determinism is compatible with being a free agent. You endorse a materialist philosophy of freedom as well. An intellectually honest person open to empirical data, you simply believe that something along these lines must be true.
Now you try to feel that it is true. You try to consciously experience the fact that at any given moment of your life, you could not have acted otherwise. You try to experience the fact that even your thoughts, however rational and moral, are predetermined — by something unconscious, by something you can not see. And in doing so, you start fooling around with the conscious self-model Mother Nature evolved for you with so much care and precision over millions of years: You are scratching at the user-surface of your own brain, tweaking the mouse-pointer, introspectively trying to penetrate into the operating system, attempting to make the invisible visible. You are challenging the integrity of your phenomenal self by trying to integrate your new beliefs, the neuroscientific image of man, with your most intimate, inner way of experiencing yourself. How does it feel?
I think that the irritation and deep sense of resentment surrounding public debates on the freedom of the will actually has nothing much to do with the actual options on the table. It has to do with the perfectly sensible intuition that our presently obvious answer will not only be emotionally disturbing, but ultimately impossible to integrate into our conscious self-models.
Or our societies: The robust conscious experience of free will also is a social institution, because the attribution of accountability, responsibility, etc. are the decisive building blocks for modern, open societies. And the currently obvious answer might be interpreted by many as having clearly anti-democratic implications: Making a complex society work implies controlling the behavior of millions of people; if individual human beings can control their own behavior to a much lesser degree than we have thought in the past, if bottom-up doesn't work, then it becomes tempting to control it top-down, by the state. And this is the second way in which enlightenment could devour its own children. Yes, free will truly is a dangerous question, but for different reasons than most people think. "
Our commonsense conceptions of ourselves have co-evolved over hundreds of thousands of years, along with their physiological, homeostatic, neuroendocrine, and limbic emotional correlates. This whole complex (us, that is) can be upset by facing what it can come to know to be true about the impersonal physical processes that actually run our show, finding it impossible to integrate its 'illusory' self image.
Here is a clip, and then its more extended context from the piece by Metzinger on edge.org..his response to the question "What is your dangerous idea." He frames it much better than I can. First the clip:
"I think that the irritation and deep sense of resentment surrounding public debates on the freedom of the will actually has nothing much to do with the actual options on the table. It has to do with the perfectly sensible intuition that our presently obvious answer will not only be emotionally disturbing, but ultimately impossible to integrate into our conscious self-models."
Then the more extended quotation:
"For middle-sized objects at 37° like the human brain and the human body, determinism is obviously true. The next state of the physical universe is always determined by the previous state. And given a certain brain-state plus an environment you could never have acted otherwise. A surprisingly large majority of experts in the free-will debate today accept this obvious fact...."
"Yes, you are a physically determined system. But this is not a big problem, because, under certain conditions, we may still continue to say that you are "free": all that matters is that your actions are caused by the right kinds of brain processes and that they originate in you. A physically determined system can well be sensitive to reasons and to rational arguments, to moral considerations, to questions of value and ethics, as long as all of this is appropriately wired into its brain. You can be rational, and you can be moral, as long as your brain is physically determined in the right way. You like this basic idea: physical determinism is compatible with being a free agent. You endorse a materialist philosophy of freedom as well. An intellectually honest person open to empirical data, you simply believe that something along these lines must be true.
Now you try to feel that it is true. You try to consciously experience the fact that at any given moment of your life, you could not have acted otherwise. You try to experience the fact that even your thoughts, however rational and moral, are predetermined — by something unconscious, by something you can not see. And in doing so, you start fooling around with the conscious self-model Mother Nature evolved for you with so much care and precision over millions of years: You are scratching at the user-surface of your own brain, tweaking the mouse-pointer, introspectively trying to penetrate into the operating system, attempting to make the invisible visible. You are challenging the integrity of your phenomenal self by trying to integrate your new beliefs, the neuroscientific image of man, with your most intimate, inner way of experiencing yourself. How does it feel?
I think that the irritation and deep sense of resentment surrounding public debates on the freedom of the will actually has nothing much to do with the actual options on the table. It has to do with the perfectly sensible intuition that our presently obvious answer will not only be emotionally disturbing, but ultimately impossible to integrate into our conscious self-models.
Or our societies: The robust conscious experience of free will also is a social institution, because the attribution of accountability, responsibility, etc. are the decisive building blocks for modern, open societies. And the currently obvious answer might be interpreted by many as having clearly anti-democratic implications: Making a complex society work implies controlling the behavior of millions of people; if individual human beings can control their own behavior to a much lesser degree than we have thought in the past, if bottom-up doesn't work, then it becomes tempting to control it top-down, by the state. And this is the second way in which enlightenment could devour its own children. Yes, free will truly is a dangerous question, but for different reasons than most people think. "
Subscribe to:
Posts (Atom)