Tsuchiya and Adolphs at Cal. Tech. offer a broad review of this topic in Trends in Cognitive Sciences (Volume 11, Issue 4, April 2007, Pages 158-167) which has some excellent illustrations for teaching. Here I show the abstract and three figures from the article, the PDF is
HERE.

Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html).

Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].

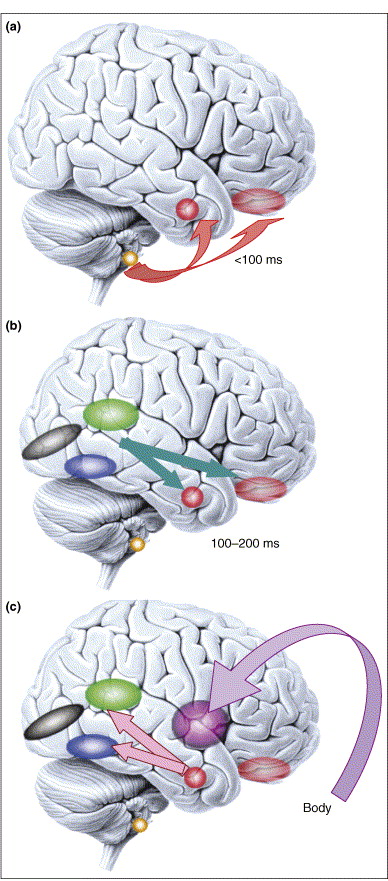
Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].