After the Chopin B minor Scherzo...
 .....Deric with his first (1969, Ann Gordon Walker) and last (1995, Dima Klenchin) postdoctoral students
.....Deric with his first (1969, Ann Gordon Walker) and last (1995, Dima Klenchin) postdoctoral students
.....Deric with his first (1969, Ann Gordon Walker) and last (1995, Dima Klenchin) postdoctoral students
.....Deric with his first (1969, Ann Gordon Walker) and last (1995, Dima Klenchin) postdoctoral students
Face perception elicits activation within a distributed cortical network in the human brain. The network includes visual (‘‘core’’) regions, as well as limbic and prefrontal (‘‘extended’’) regions, which process invariant facial features and changeable aspects of faces, respectively. We used functional Magnetic Resonance Imaging and Dynamic Causal Modeling to investigate effective connectivity and functional organization between and within the core and the extended systems. We predicted a ventral rather than dorsal connection between the core and the extended systems during face viewing and tested whether valence and fame would alter functional coupling within the network. We found that the core system is hierarchically organized in a predominantly feedforward fashion, and that the fusiform gyrus (FG) exerts the
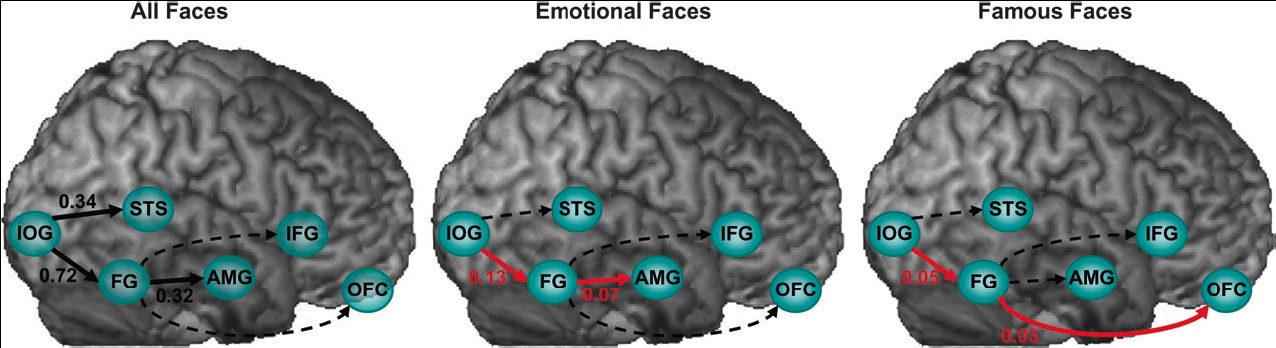
dominant influence on the extended system. Moreover, emotional faces increased the coupling between the FG and the amygdala, whereas famous faces increased the coupling between the FG and the orbitofrontal cortex. Our results demonstrate content-specific dynamic alterations in the functional coupling between visuallimbic and visual-prefrontal face-responsive pathways.Face perception elicits activation within a distributed cortical network. Axial sections, taken from a representative subject, illustrate activation within the core (IOG-inferior occipital gyrus, FG-fusiform gyrus, STS - superior temporal sulcus) and extended (AMG-amygdala, IFG-inferior frontal gyrus, OFC-orbitofrontal cortex) systems. Coordinates are in the Talaraich space. L 5 left, R 5 right.
(click to enlarge) Alterations in effective connectivity within the core and the extended systems induced by all faces, emotional faces, and famous faces. Black connections indicate significant regional effects, red connections indicate significant bilinear effects, and dotted lines indicate non-significant effects.
...shows that amygdala responses depend on visual awareness. Under conditions in which subjects were not aware of fearful faces flashed for 33 ms, no differential activation was observed in the amygdala. On the other hand, differential activation was observed for 67 ms fearful targets that the subjects could reliably detect. When trials were divided into hits, misses, correct rejects, and false alarms, we show that target visibility is an important factor in determining amygdala responses to fearful faces. Taken together, our results further challenge the view that amygdala responses occur automatically.Duncan and Barrett, in their commentary, suggest
...that the amygdala is acting to increase neural activity in the fusiform gyrus, thereby increasing the likelihood that visual representations that have affective value reach awareness. The psychological consequence is that a person’s momentary affective state might help to select the contents of conscious experience.Visual awareness is associated with amygdala activation. In the Pessoa et al. study, participants viewed backwardly masked images of faces that depicted fear, presented for either 33 ms or 67 ms. All participants showed greater amygdala activation when viewing fearful faces that were presented for 67 ms, compared with faces that depicted neutral expressions. (a) Pessoa et al. found an increase in amygdala activation (as well as fusiform gyrus activation, which is not shown in the figure) only among those participants who showed objective awareness of 33 ms presentations of faces that depicted fear. (b) Participants who did not show objective awareness did not have significant increases in amygdala activation. Given the excitatory projections from the amygdala to the ventral visual stream, this finding suggests that the amygdala enhances visual awareness for objectives with affective value.
challenges the conventional models of human perception, which assume that sensory experience involves a critical interplay between activity in sensory cortex representing the stimulus and pre-frontal cortex which serves as an “observer” system- receiving and interpreting the patterns of activity originating in posterior, sensory cortex.In an fMRI study in which individuals were exposed to a highly engaging popular movie they find:
a surprisingly robust and wide-spread activation of most of the posterior part of the brain- which was remarkably “synchronized” across individuals watching the same movie. These results attest to the massive engagement of sensory cortex by naturalistic sensory stimuli. However, in contrast to this wide-spread activation in sensory cortex- we have found a remarkably little activation in frontal areas of the brain (Fig. 1).Fig. 1 Brain activation during repeated movie presentation. Yellow regions show highly activated areas in posterior, sensory cortex. Note lack of activation in frontal areas and in the intriguing “intrinsic” islands in posterior cortex (arrows).
we have conducted a series of experiments in which we mapped brain activity during tasks that were explicitly targeted at eliciting “self-related” brain activity- such as introspection to visual and auditory modalities, or self-judgment evaluations. Although these were high level cognitive tasks of extreme complexity- our results show a remarkably consistent pattern of activity focused primarily on pre-frontal cortex (see yellow patches in figure 2)...The critical question was now- how do these areas behave when the task shifts to a purely perceptual one? The results were clear cut- the activity shifted now to a different set of areas- located more posteriorly in sensorymotorThe results are:
cortex (green patches in fig. 2).Fig. 2 Comparing brain activity during reflective self-related tasks and during intense perception. Prefrontal areas (yellow patches) showed significant activation during self-related introspection and self judgment tasks. However a completely different and highly segregated network of more posterior areas were active during engaging perceptual tasks (green patches). No overlap was found between the two networks, and the prefrontal cortex was actually inhibited during intense perception. The results clearly rule-out the need for an “observer” function in self-related
cortex during perception.
... actually compatible with the strong intuitive sense we have of 'losing our selves' in a highly engaging sensory-motor act (such as watching a movie). They are intriguingly reminiscent of recurrent eastern philosophical themes which emphasize the ‘silencing' of the self during intense engagement with the outside world.
The implications of the discovery of mirroring mechanisms and embodied simulation for empathetic responses to images in general, and to works of visual art in particular, have not yet been assessed. Here, we address this issue and we challenge the primacy of cognition in responses to art. We propose that a crucial element of esthetic response consists of the activation of embodied mechanisms encompassing the simulation of actions, emotions and corporeal sensation, and that these mechanisms are universal. This basic level of reaction to images is essential to understanding the effectiveness both of everyday images and of works of art. Historical, cultural and other contextual factors do not preclude the importance of considering the neural processes that arise in the empathetic understanding of visual artworks.

Figure 1. Embodied simulation in esthetic experience: actions. The sense of exertion, which Michelangelo intended his figures to show as they struggle to escape from the block of stone, is effectively conveyed to the spectator. Michelangelo, Slave called Atlas, Florence, Academia (ca. 1520–1523), marble © Scala/Art Resource, NY.

Figure 3. Embodied simulation in esthetic experience: implied gestures of the artist. The gestures that are only implicit in the marks on these works of art are corporeally felt by their spectators. (a) Jackson Pollock, Number 14: Gray (1948), enamel over gesso on paper, Yale University Gallery, The Katharine Ordway Collection © 2004 The Pollock-Krasner Foundation/Artists Rights Society (ARS), New York. (b) Lucio Fontana, Concetto Spaziale ‘Atteza’ (‘Waiting’) (1960), canvas, Tate Gallery, London © Tate Gallery, London/Art Resource, NY/ Fondazione Lucio Fontana, Milano.

Figure 4. The mirror neuron system in monkeys and humans. (a) Activation of the area F5 mirror neuron during motor-act observation. (b) Activation of the area F5 mirror neuron during action execution. For both conditions, six consecutive rasters (spike recordings) during six consecutive trials are shown. The arrows indicate the onset of observed and executed grasping. (a) and (b) modified, with permission, from Ref. [60]. (c) Somatotopy of premotor and parietal cortices as revealed by fMRI during action observation. Activation foci are projected on the lateral surface of a standard brain (MNI). Red, activation during the observation of mouth grasping; green, activation during the observation of hand grasping; blue, activation during the observation of foot kicking. Overlap of colors indicates activation foci present during observation of actions made by different effectors.


The hippocampus plays a time-limited role in the formation of declarative memories, with memories gradually becoming independent of the hippocampus over time. It is believed that these remote memories are gradually transferred from the hippocampus to the neocortex for long-term storage..investigations indicate that the medial prefrontal cortex (mPFC) is critical for the consolidation of hippocampal-dependent memories. In trace-conditioning tasks for instance, hippocampal lesions cause a severe deficit when made soon after training, but not after a month, whereas mPFC lesions produce the opposite pattern of impairments. ...The role of mPFC activity in memory formation remains unclear. One possibility is that mPFC affects the transfer of hippocampal activity toward the neocortex. Consistent with this possibility, the mPFC projects to the rhinal cortices, the main route for impulse traffic into and out of the hippocampus...the present study was undertaken to test the idea that the mPFC influences memory formation by modulating interactions between the neocortex and hippocampus at the level of the rhinal cortices. To this end, we examined the relative timing of unit activity in the mPFC, PR, and ER cortices during the acquisition of a trace-conditioning task.And, here is the abstract of the paper:
Much data suggests that hippocampal–medial prefrontal cortex (mPFC) interactions support memory consolidation. This process is thought to involve the gradual transfer of transient hippocampal-dependent memories to distributed neocortical sites for long-term storage. However, hippocampal projections to the neocortex involve a multisynaptic pathway that sequentially progresses through the entorhinal and perirhinal regions before reaching the neocortex. Similarly, the mPFC influences the hippocampus via the rhinal cortices, suggesting that the rhinal cortices occupy a strategic position in this network. The present study thus tested the idea that the mPFC supports memory by facilitating the transfer of hippocampal activity to the neocortex via an enhancement of entorhinal to perirhinal communication. To this end, we simultaneously recorded mPFC, perirhinal, and entorhinal neurons during the acquisition of a trace-conditioning task in which a visual conditioned stimulus (CS) was followed by a delay period after which a liquid reward was administered. At learning onset, correlated perirhinal-entorhinal firing increased in relation to mPFC activity, but with no preferential directionality, and only after reward delivery. However, as learning progressed across days, mPFC activity gradually enhanced rhinal correlations in relation to the CS as well, and did so in a specific direction: from entorhinal to perirhinal neurons. This suggests that, at late stages of learning, mPFC activity facilitates entorhinal to perirhinal communication. Because this connection is a necessary step for the transfer of hippocampal activity to the neocortex, our results suggest that the mPFC is involved in the slow iterative process supporting the integration of hippocampal-dependent memories into neocortical networks.
Estrogens are classically viewed as hormones that bind to intracellular receptors, which then act as transcription factors to modulate gene expression; however, they also affect many aspects of neuronal functioning by rapid nongenomic actions. Brain estrogen production can be regulated within minutes by changes in aromatase (estrogen synthase) activity as a result of calcium-dependent phosphorylations of the enzyme. To determine the effects of rapid changes in estrogen availability on male copulatory behavior, we mimicked in male mice the rapid upregulation and downregulation of brain estrogen concentration that should occur after inactivation or activation of aromatase activity. A single injection of different aromatase inhibitors [Vorozole, 1,4,6-androstatrien-3,17-dione (ATD), or its metabolite 17-OH-ATD (1,4,6-androstatrien-17ß-ol-3-one)] almost completely suppressed male sexual behavior (mounts and intromissions) expressed 10–20 min later by C57BL/6J mice but did not affect behavior in aromatase knock-out (ArKO) mice, activated by daily injections of estradiol benzoate, thereby confirming the specificity of the behavioral inhibition observed in wild-type mice. The rapid ATD-induced inhibition was reversed by the simultaneous injection of a large dose of estradiol. A single injection of estradiol to ArKO mice also activated male sexual behavior within 15 min. Thus, rapid increases or decreases in brain estrogen concentrations are followed within minutes by corresponding changes in male sexual behavior. Sexual behavior can thus be used to monitor changes in local estrogen concentrations and analyze the mechanisms mediating the rapid decline in estrogen signaling that takes place after inhibition of estrogen synthesis.

Systems neuroscience aims to understand how billions of neurons in the mammalian brain support goal-directed behavior, such as decision making. Deciphering how individual neurons respond to sensory inputs or motor decisions has focused on delineating the neural basis of these processes in discrete regions of the brain's cortex, and has provided key insights into the physiological basis of behavior. However, evidence from neuropsychological, electrophysiological, and neuroimaging studies in humans has revealed that interactions between widespread neural regions in the brain underlie fluid, organized behavior.He then summarizes work reported in three papers to:
...unravel the details of these interactions by assessing the simultaneous activity of neurons in multiple sites of the mammalian brain. The studies show that network interactions among anatomically discrete brain regions underlie cognitive processing and dispel any phrenological notion that a given innate mental faculty is based solely in just one part of the brain.
...Taken together, the three papers indicate that top-down signals between brain regions regulate the flow of information and that distributed neural networks that use oscillatory dynamics support a broad spectrum of neural processing and behavior. The results in cats and monkeys also nicely parallel findings in humans...this particular coupling mechanism is used to delineate task-specific network activity

Civil societies function because people pay taxes and make charitable contributions to provide public goods. One possible motive for charitable contributions, called "pure altruism," is satisfied by increases in the public good no matter the source or intent. Another possible motive, "warm glow," is only fulfilled by an individual's own voluntary donations. Consistent with pure altruism, we find that even mandatory, tax-like transfers to a charity elicit neural activity in areas linked to reward processing. Moreover, neural responses to the charity's financial gains predict voluntary giving. However, consistent with warm glow, neural activity further increases when people make transfers voluntarily. Both pure altruism and warm-glow motives appear to determine the hedonic consequences of financial transfers to the public good.Figure: Neural response in the ventral striatum to mandatory payoffs for the subject (yellow), the charity (blue), and both (green). (To test for the pure altruism and warm-glow motives, they used functional magnetic resonance imaging while subjects played a dictator game. Subjects received $100 and then made decisions about whether or not to give money to a local food bank. They also observed mandatory, tax-like transfers of their money to the food bank.)
They suggest that:
...This result supports arguments for a common "neural currency" of reward and shows that this model can be applied not just to choice over money, risk, and private consumption goods, but also to more abstract policy choices involving taxation and charitable giving. Our results are also important for understanding why people give money to charitable organizations. First, these transfers are associated with neural activation similar to that which comes from receiving money for oneself. The fact that mandatory transfers to a charity elicit activity in reward-related areas suggests that even mandatory taxation can produce satisfaction for taxpayers. A better understanding of the conditions under which taxation elicits "neural rewards" could prove useful for evaluating the desirability of different tax policies.
Although the experience of disgust feels primal, the emotion does not seem to be widespread in other animals. Many species exhibit distaste in response to the sensory properties of food — such as sourness and bitterness — and a monkey, cat or human infant might spit out something disagreeable. But only humans beyond infancy will reject food on the basis of where it might have been and what it might have touched.
A clue is the language of moral indignation itself...All cultures and languages that we have studied have at least one word that applies both to core disgust (cockroaches and faeces) and also to some kind of social offence, such as sleazy politicians or hypocrites...people labelled as disgusting in this way evoke fears of contamination just as rotting food does...disgust drives some moral judgements, but ... they are mainly those relating to behaviour that involves bodily fluids or contact — gay sex, for instance — rather than more abstract issues.
Clues suggest a physiological reality for moral disgust. Whereas anger pushes the heart rate up, being viscerally disgusted makes it drop. Experiments done by Haidt and Sherman showed
... people hooked up to a heart monitor video footage of morally negative but not viscerally disgusting behaviour, such as an American neo-Nazi meeting. The participants said that the video triggered disgust and anger, and on average their heart rates fell, not rose. What's more, those who reported increased clenching in their throat had a greater drop in heart rate, making the link with core disgust look stronger.
...this is the first physiological evidence that socio-moral disgust really is disgust and not just metaphor or anger.
Brain imaging studies might also point to an overlap between core and moral disgust... Moll...used magnetic resonance imaging (MRI) to monitor the flow of blood in the brains of 13 healthy adult volunteers as they mulled over situations evocative of core disgust and those that elicit self-reported moral disgust or indignation. He found that core and moral disgust recruit overlapping brain areas, particularly the lateral and medial orbitofrontal cortex, suggesting that the emotions are related. These regions of the brain are activated by unpleasant sensory stimuli, and they connect with other emotion-related areas, such as the amygdala.
Evolution suggests that the human moral faculty — the psychological systems that make judgements about right and wrong, what's permissible and what isn't — was cobbled together from pre-existing brain systems over millions of years of biological and cultural evolution. Along the way, it latched onto disgust as a useful tool...The experimental data point to the possibility that our disgust system might have been adapted by evolution to allow us to reject or disapprove of abstract concepts such as ideologies and political views that are deeply influenced by culture, as well social groups associated with 'disgusting' concepts...In making symbolic distinctions between us and them visceral, disgust could potentially foster greater cohesion within groups by bringing people together in defence against a common out-group...Disgust works for the group as it does for the individual — what is in the group is 'me' and what is not is 'not me'...Where core disgust is the guardian of the body, moral disgust acts as the guardian of social body — that's when disgust shows its ugliest side....disgust is an emotion we are stuck with. Heuser suggests that the challenge... is to make people more reflective about what they say and think. He cites the success that advocates of political correctness have had in lowering the prevalence of casually sexist and racist language. Moll suggests optimistically that cultivating cultural and personal values of tolerance and empathy could function as an antidote to the toxic effects of disgust...by thinking less with our guts, and more with our heads and hearts, we might be able push back the boundaries of our moral world.