Valse Romantique by Debussy, on my Steinway B at Twin Valley...
Monday, July 09, 2007
Ambiguity and anxiety: overreaction and a serotonin receptor
Nader and Alleine describe work of Tsetsenis et al.
Here is the abstract from Tsetsenis et al.
When the threat of terrorist attack is elevated, the United States Department of Homeland Security changes its prediction of danger from yellow to orange to red. Most of us can manage our levels of vigilance and of anxiety appropriately in response to these cues. However, imagine how debilitating it would be if you were unable to manage your anxiety and reduce your fear of attack when the threat level was reduced. In fact, many people with anxiety disorders suffer from precisely this kind of condition. The paper by Tsetsenis et al. finds that mice lacking the serotonin 1a receptor overreact to ambiguous predictors of aversive events in this same way, providing insight into factors that could predispose individuals to such disorders and into the neural locus of the effect.
Understanding the neural bases of contingency is not simply an academic question, but very much a mental health one. Unpredictable aversive events can be much more stressful than the same events when they are anticipated. In addition, psychopathologies can influence the perception of contingency. For example, depressed people have a different sense than nondepressed people of how their responses affect the environment, and exposure to unpredictable or uncontrollable aversive events is suggested to directly influence the development of depression.
The new study by Tsetsenis et al. makes a significant contribution to our understanding of the neural mechanisms mediating contingency learning. The authors studied mice in which the serotonin 1a receptor (Ht1a) gene was knocked out or inactivated during development. This receptor causes membrane hyperpolarization of nonserotonergic neurons and acts as an autoreceptor on serotonergic neurons in the raphe. Ht1a dysfunction is linked to anxiety disorders and depression, and mice lacking Ht1a receptors show increased avoidance behavior. This phenotype is attributable to the absence of the Ht1a in the forebrain during development; eliminating these receptors during adulthood does not cause the mice to show the anxious phenotype. Although fear of an aversive context is comparable in knockout and wild-type mice, the knockout mice over-generalize their fear of the 'aversive' context to a similar context containing novel elements, a situation in which wild-type mice are able to decrease their fear levels11. This finding suggested that these mutant mice focus unduly on cues that have been paired with shock rather than on cues that have not.Figure: Humans and other animals can accurately estimate the probability of danger from their experience of specific environments or cues and use this information to respond appropriately. A normal mouse (top) accurately estimates the threat from an ambiguous cue, a sleeping cat, and a less ambiguous cue, an alert cat, and is appropriately cautious or alarmed, respectively. In contrast, anxious people and animals, such as the Htr1a knockout mouse assessed by Tsetsenis et al. (bottom), often overestimate the danger represented by ambiguous cues and over-respond, given the level of threat. This is likely to interfere with the need to respond to other important events in the environment (such as cheese).

Here is the abstract from Tsetsenis et al.
Serotonin receptor 1A knockout (Htr1aKO) mice show increased anxiety-related behavior in tests measuring innate avoidance. Here we demonstrate that Htr1aKO mice show enhanced fear conditioning to ambiguous conditioned stimuli, a hallmark of human anxiety. To examine the involvement of specific forebrain circuits in this phenotype, we developed a pharmacogenetic technique for the rapid tissue- and cell type–specific silencing of neural activity in vivo. Inhibition of neurons in the central nucleus of the amygdala suppressed conditioned responses to both ambiguous and nonambiguous cues. In contrast, inhibition of hippocampal dentate gyrus granule cells selectively suppressed conditioned responses to ambiguous cues and reversed the knockout phenotype. These data demonstrate that Htr1aKO mice have a bias in the processing of threatening cues that is moderated by hippocampal mossy-fiber circuits, and suggest that the hippocampus is important in the response to ambiguous aversive stimuli.
Friday, July 06, 2007
Threads of our lives in dreams...
 Rebecca Cathcart writes an article on dreaming and "big dreams" (PDF here) that resonates with my own experience of having, particularly before waking, emotionally intense dreams whose story line seems to be an obvious attempt to integrate important personal issues. Some clips:
Rebecca Cathcart writes an article on dreaming and "big dreams" (PDF here) that resonates with my own experience of having, particularly before waking, emotionally intense dreams whose story line seems to be an obvious attempt to integrate important personal issues. Some clips:Big dreams are once again on the minds of psychologists as part of a larger trend toward studying dreams as meaningful representations of our concerns and emotions...The dreaming imagination does not just harvest images from remembered experience...It has a “poetic creativity” that connects the dots and “deforms the given,” turning scattered memories and emotions into vivid, experiential vignettes that can help us to reflect on our lives....Cultural narratives in regions like Vietnam and North and South America assign special importance to such dreams and consider them actual encounters with the spirits of lost loved ones...This notion is so widely shared by traditions all across the globe that some scholars have gone so far as to argue that religion itself actually originated in dream experience.The article emphasizes the role of dreams in dealing with death and grief.
Grief itself is transformative. It is a process of disassembly. The bereaved must let go of the selves they were, as well as the loved ones they have lost. The dreams we have while grieving are an important part of that process...Our dreams have to do with how we internalize the people we love...You learn to look within for the loved one and the particular function that person played in your life, such as caretaking or guidance in the case of a parent. This becomes part of a function that you can provide for yourself.
Dreams that occur during rapid eye movement, or REM, cycles are the most memorable and emotionally powerful...The dreams have power because brain activity during REM is most similar to that of a waking state. The emotional responses to REM dream content, therefore, are most like the responses during waking cognition...Core body temperature rises gradually from its nadir in the middle of the night during slow-wave sleep, the least active brain state. As morning nears, subcortical brain activity tied to the circadian cycle increases. When these cycles coincide in the last and longest REM phase... the mind produces its most dramatic dreams...the four or five phases of REM in a normal night’s sleep might include similar dream content. Just as the image of a lost loved one stimulates parts of the brain associated with loss, the content of dreams early in the sleep cycle could set the tone for that night’s dream experiences. Our memories upon waking, therefore, may be our recollection of a night’s cumulative dream content.
Thursday, July 05, 2007
Where the brain understands animate agents..
Wheatley et al offer an interesting study in a recent issue of Psychological Science (vol 18, pg 469, 2007, PDF here). Here is the abstract and two figures:
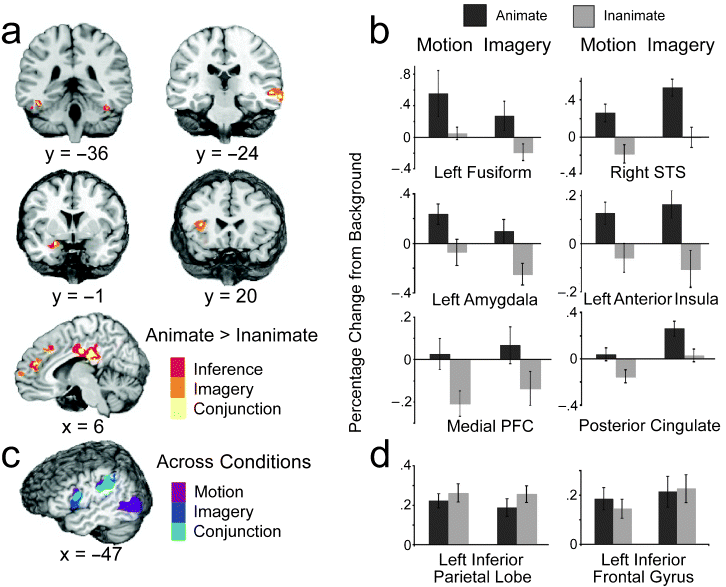
How people understand the actions of animate agents has been vigorously debated. This debate has centered on two hypotheses focused on anatomically distinct neural substrates: The mirror-system hypothesis proposes that the understanding of others is achieved via action simulation, and the social-network hypothesis proposes that such understanding is achieved via the integration of critical biological properties (e.g., faces, affect). In this study, we assessed the areas of the brain that were engaged when people interpreted and imagined moving shapes as animate or inanimate. Although observing and imagining the moving shapes engaged the mirror system, only activation of the social network was modulated by animacy.Lateral and medial views of the social network (top, highlighted in yellow) and mirror system (bottom, highlighted in blue). The social network includes areas associated with biological motion (superior temporal sulcus, labeled "1"), biological form (lateral fusiform gyrus, labeled "6"), mentalizing (medial prefrontal cortex and posterior cingulate, labeled "3" and "4," respectively), and affective processing (insula and amygdala, labeled "2" and "5," respectively). The mirror system consists of the inferior parietal cortex (labeled "7") and the ventral-premotor/inferior-frontal cortex (labeled "8").
Experimental results. The brain slices in (a) depict areas of the social network that were more active when moving shapes were inferred (red) or imagined (orange) as animate than when they were inferred or imagined as inanimate. Yellow areas were more active for both animate inference and imagery ("conjunction"). The graph in (b) displays the average hemodynamic responses within the conjunction areas as a function of animacy (animate, inanimate) and condition (motion, imagery). (Results are not shown for the posterior insula, although this was also a conjunction area.) The illustration in (c) shows areas of the mirror system that were more active when subjects watched and made inferences about the moving shapes (purple) and when they imagined (dark blue) the moving shapes relative to when they viewed the backgrounds alone; light-blue areas were more active during both the motion and imagery conditions ("conjunction") than in the background condition. The graph in (d) shows the average hemodynamic responses of the conjunction mirror areas as a function of animacy and condition. For purposes of illustration, all group data are presented on the N27 (AFNI software) brain. Error bars represent standard errors. STS = superior temporal sulcus; PFC = prefrontal cortex.

Yawn to cool your brain?
 A curious and slightly flakey bit: Eric Nagourney in Tuesday's Science section of the NY Times describes work by Gallup et al (PDF here) published in the Journal Evolutionary Psychology. It seems to me they might have actually measured brain temperature instead of just speculating about it. Nagourney notes the proposal by Gallup et al. that:
A curious and slightly flakey bit: Eric Nagourney in Tuesday's Science section of the NY Times describes work by Gallup et al (PDF here) published in the Journal Evolutionary Psychology. It seems to me they might have actually measured brain temperature instead of just speculating about it. Nagourney notes the proposal by Gallup et al. that:yawning... is a way for the body to cool the brain...volunteers yawned more often in situations in which their brains were likely to be warmer...To prove their theory that yawning regulates brain temperature when other systems in the body are not doing enough, the researchers took advantage of the well-established tendency of people to yawn when those around them do — the so-called contagious yawn...The volunteers were asked to step into a room by themselves and watch a video showing people behaving neutrally, laughing or yawning. Observers watching through a one-way mirror counted how many times the volunteers yawned...Some volunteers were asked to breathe only through their noses as they watched. Later, volunteers were asked to press warm or cold packs on their foreheads...“The two conditions thought to promote brain cooling (nasal breathing and forehead cooling) practically eliminated contagious yawning,” the researchers wrote.
The study may also help explain why yawning spreads from person to person...A cooler brain, Dr. Gallup said, is a clearer brain...So yawning actually appears to be a way to stay more alert. And contagious yawning, he said, may have evolved to help groups remain vigilant against danger.
Wednesday, July 04, 2007
Is Darwin due for an upgrade?
 I would recommend you have a look at this article by Douglas Erwin (PDF here) on the prospects of a paradigm shift in evolutionary biology. Here are some clips:
I would recommend you have a look at this article by Douglas Erwin (PDF here) on the prospects of a paradigm shift in evolutionary biology. Here are some clips:...constructed from the 1930s to 1950s by early geneticists, paleontologists and others..the modern synthesis...holds that mutations to DNA create new variants of existing genes within a species. Natural selection, driven by competition for resources, allows the best-adapted individuals to produce the most surviving offspring. So adaptive variants of genes become more common...Computer simulations have shown how selection can produce a complex eye from a simple eyespot in just a few hundred thousand years.
In the past few years every element of this paradigm has been attacked. Concerns about the sources of evolutionary innovation and discoveries about how DNA evolves have led some to propose that mutations, not selection, drive much of evolution, or at least the main episodes of innovation, like the origin of major animal groups, including vertebrates.
Comparative studies of development have illuminated how genes operate, and evolve, and this places less emphasis on the gradual accumulation of small genetic changes emphasized by the modern synthesis. Work in ecology has emphasized the role organisms play in building their own environments, and studies of the fossil record raise questions about the role of competition. The last major challenge has argued for a hierarchical view of evolution, with selection occurring at many levels, including between species.
The Achilles’ heel of the modern synthesis.. is that it deals primarily with the transmission of genes from one generation to the next, but not how genes produce bodies. The recent discoveries in the new field of evolutionary developmental biology, or evo-devo, that the gene Pax-6 controls the formation of eyes in mice and humans, Nkx2.5 heart formation, and a suite of other genes the formation of the nervous system, has provided a means to investigate the genetic and developmental mechanisms influencing how the form of organisms has evolved, not just their genes.
Core gene networks appeared long ago that locked development onto a certain path (an example being a kernel of five key genes regulating development of the gut that appeared 500 million years ago). These events, small and large, limit the range of possibilities on which natural selection can act...just as the erosive power of a river changes the future options for the course of the river, so evolution itself changes future evolutionary possibilities.
The first cyanobacteria turned carbon dioxide into oxygen and set off a revolution that completely changed the chemistry of the oceans and atmosphere. Most species modify their environment and this often changes how selection affects them: they construct, at least in part, their own environment. As evolutionary biologists we have little understanding of what these processes mean for evolution.
Does all this add up to a new modern synthesis? There is certainly no consensus among evolutionary biologists, but development, ecology, genetics and paleontology all provide new perspectives on how evolution operates, and how we should study it. None of these concerns provide a scintilla of hope for creationists, as scientific investigations are already providing new insights into these issues. The foundations for a paradigm shift may be in place, but it may be some time before we see whether a truly novel perspective develops or these tensions are accommodated within an expanded modern synthesis.
Tuesday, July 03, 2007
A social/musical at Twin Valley
Every year or two my partner Len and I host a social afternoon on a Sunday from 3-6 at our home on Twin Valley Road in the Town of Middleton, Wisconsin. It is a stone schoolhouse built in 1860, converted to a residence by a Frank Lloyd Wright lieutenant named Herb Fritz. Len (who is a gourmet cook) provides appetizers and Hors D'oeuvres and I do some piano playing either by myself or with a string chamber group. Here are two pictures from Sunday July 1. (By the way, blog posts are going to be spotty this holiday week).

After the Chopin B minor Scherzo...
 .....Deric with his first (1969, Ann Gordon Walker) and last (1995, Dima Klenchin) postdoctoral students
.....Deric with his first (1969, Ann Gordon Walker) and last (1995, Dima Klenchin) postdoctoral students

After the Chopin B minor Scherzo...
 .....Deric with his first (1969, Ann Gordon Walker) and last (1995, Dima Klenchin) postdoctoral students
.....Deric with his first (1969, Ann Gordon Walker) and last (1995, Dima Klenchin) postdoctoral students
Science of the Soul
I recommend this engaging article by Cornelia Dean (PDF here) on how evolutionary biologists and cognitive scientists are discovering physical bases for the feelings from which moral sense emerges — not just in people but in other animals as well.
As is illustrated by the three Republican presidential candidates that raised their hands during a debate when asked 'Is there anyone who does not believe in evolution?,' challenges to the uniqueness of humanity in creation are just as alarming as the Copernican assertion that Earth is not the center of the universe,
As is illustrated by the three Republican presidential candidates that raised their hands during a debate when asked 'Is there anyone who does not believe in evolution?,' challenges to the uniqueness of humanity in creation are just as alarming as the Copernican assertion that Earth is not the center of the universe,
Monday, July 02, 2007
Face perception by distributed cortical networks.
Continuing to pass along material from talks given at the recent ASSC meeting, here is the abstract and some figures from an interesting bit of work on face perception (PDF here).
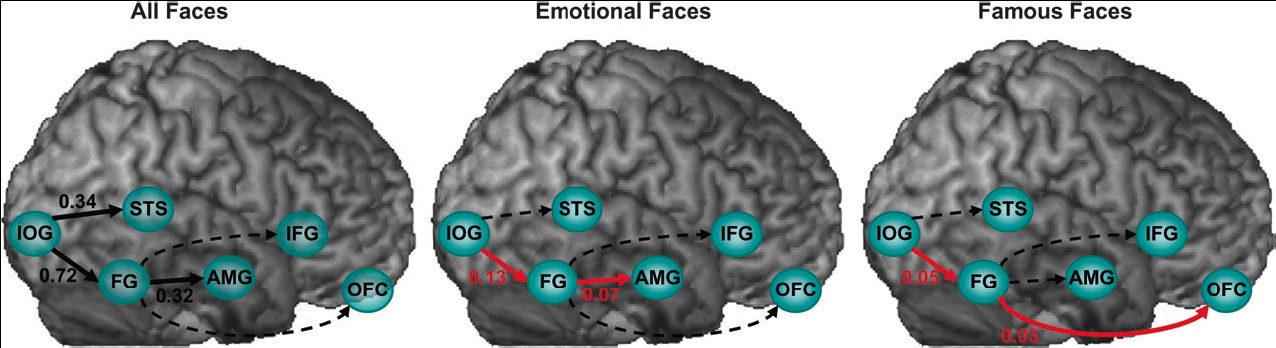
Face perception elicits activation within a distributed cortical network in the human brain. The network includes visual (‘‘core’’) regions, as well as limbic and prefrontal (‘‘extended’’) regions, which process invariant facial features and changeable aspects of faces, respectively. We used functional Magnetic Resonance Imaging and Dynamic Causal Modeling to investigate effective connectivity and functional organization between and within the core and the extended systems. We predicted a ventral rather than dorsal connection between the core and the extended systems during face viewing and tested whether valence and fame would alter functional coupling within the network. We found that the core system is hierarchically organized in a predominantly feedforward fashion, and that the fusiform gyrus (FG) exerts the
dominant influence on the extended system. Moreover, emotional faces increased the coupling between the FG and the amygdala, whereas famous faces increased the coupling between the FG and the orbitofrontal cortex. Our results demonstrate content-specific dynamic alterations in the functional coupling between visuallimbic and visual-prefrontal face-responsive pathways.Face perception elicits activation within a distributed cortical network. Axial sections, taken from a representative subject, illustrate activation within the core (IOG-inferior occipital gyrus, FG-fusiform gyrus, STS - superior temporal sulcus) and extended (AMG-amygdala, IFG-inferior frontal gyrus, OFC-orbitofrontal cortex) systems. Coordinates are in the Talaraich space. L 5 left, R 5 right.
(click to enlarge) Alterations in effective connectivity within the core and the extended systems induced by all faces, emotional faces, and famous faces. Black connections indicate significant regional effects, red connections indicate significant bilinear effects, and dotted lines indicate non-significant effects.

Friday, June 29, 2007
A Schubert Scherzo
....from his Sonata No. 10. I thought I might slip in another of my brief recordings, a bit of relief from the heavy neuroscience...
Role of the amygdala in visual awareness.
There have been several reports that subliminal stimuli (such as flashing a picture of an angry face for 33 msec) can activate the amygdala even though the subject is unaware of the stimulus. Pessoa presented work at the recent ASSC meeting using more rigorous criteria for behavioral performance that suggests, to the contrary, that visibility or attention is required for the expression of the effect of valence on early visual processing (even as early as the 'automatic' parallel processing in V1.) A PDF of a recent Pessoa et al. article is here, and PDF of commentary on this work by Duncan and Barrett here). This work:
...shows that amygdala responses depend on visual awareness. Under conditions in which subjects were not aware of fearful faces flashed for 33 ms, no differential activation was observed in the amygdala. On the other hand, differential activation was observed for 67 ms fearful targets that the subjects could reliably detect. When trials were divided into hits, misses, correct rejects, and false alarms, we show that target visibility is an important factor in determining amygdala responses to fearful faces. Taken together, our results further challenge the view that amygdala responses occur automatically.Duncan and Barrett, in their commentary, suggest
...that the amygdala is acting to increase neural activity in the fusiform gyrus, thereby increasing the likelihood that visual representations that have affective value reach awareness. The psychological consequence is that a person’s momentary affective state might help to select the contents of conscious experience.Visual awareness is associated with amygdala activation. In the Pessoa et al. study, participants viewed backwardly masked images of faces that depicted fear, presented for either 33 ms or 67 ms. All participants showed greater amygdala activation when viewing fearful faces that were presented for 67 ms, compared with faces that depicted neutral expressions. (a) Pessoa et al. found an increase in amygdala activation (as well as fusiform gyrus activation, which is not shown in the figure) only among those participants who showed objective awareness of 33 ms presentations of faces that depicted fear. (b) Participants who did not show objective awareness did not have significant increases in amygdala activation. Given the excitatory projections from the amygdala to the ventral visual stream, this finding suggests that the amygdala enhances visual awareness for objectives with affective value.

Thursday, June 28, 2007
Perception Without a Perceiver
Rafeal Malach gave an interesting talk at the recent ASSC meeting (PDF of article here) on work from his group that he thinks:
To examine this issue, and also to start mapping in more detail the functional organization of the various networks of pre-frontal cortex:
challenges the conventional models of human perception, which assume that sensory experience involves a critical interplay between activity in sensory cortex representing the stimulus and pre-frontal cortex which serves as an “observer” system- receiving and interpreting the patterns of activity originating in posterior, sensory cortex.In an fMRI study in which individuals were exposed to a highly engaging popular movie they find:
a surprisingly robust and wide-spread activation of most of the posterior part of the brain- which was remarkably “synchronized” across individuals watching the same movie. These results attest to the massive engagement of sensory cortex by naturalistic sensory stimuli. However, in contrast to this wide-spread activation in sensory cortex- we have found a remarkably little activation in frontal areas of the brain (Fig. 1).Fig. 1 Brain activation during repeated movie presentation. Yellow regions show highly activated areas in posterior, sensory cortex. Note lack of activation in frontal areas and in the intriguing “intrinsic” islands in posterior cortex (arrows).

To examine this issue, and also to start mapping in more detail the functional organization of the various networks of pre-frontal cortex:
we have conducted a series of experiments in which we mapped brain activity during tasks that were explicitly targeted at eliciting “self-related” brain activity- such as introspection to visual and auditory modalities, or self-judgment evaluations. Although these were high level cognitive tasks of extreme complexity- our results show a remarkably consistent pattern of activity focused primarily on pre-frontal cortex (see yellow patches in figure 2)...The critical question was now- how do these areas behave when the task shifts to a purely perceptual one? The results were clear cut- the activity shifted now to a different set of areas- located more posteriorly in sensorymotorThe results are:
cortex (green patches in fig. 2).Fig. 2 Comparing brain activity during reflective self-related tasks and during intense perception. Prefrontal areas (yellow patches) showed significant activation during self-related introspection and self judgment tasks. However a completely different and highly segregated network of more posterior areas were active during engaging perceptual tasks (green patches). No overlap was found between the two networks, and the prefrontal cortex was actually inhibited during intense perception. The results clearly rule-out the need for an “observer” function in self-related
cortex during perception.

... actually compatible with the strong intuitive sense we have of 'losing our selves' in a highly engaging sensory-motor act (such as watching a movie). They are intriguingly reminiscent of recurrent eastern philosophical themes which emphasize the ‘silencing' of the self during intense engagement with the outside world.
Wednesday, June 27, 2007
The Science of Gaydar
This is the title of an excellent article in New York Magazine, mentioned also in MindHacks.
Language without recursion...
My april 26 post dealt with a New Yorker article describing the work of Dan Everett on the language of an Amazon people which he interprets as showing humans can invent language without recursion. At edge.org there is now an interesting further conversation between Everett, Steven Pinker, and Robert Van Valin.
Tuesday, June 26, 2007
Esthetic experience and mirror neurons - motion, emotion and empathy
David Freedberg (Art History, Columbia Univ.) and Vittorio Gallese (Neuroscience, Parma) offer an interesting perspective on possible relationships between mirror neuron systems and artistic experience. Here is their abstract (and PDF of article is here.), followed by some of their illustrations.

Figure 2. Embodied simulation in esthetic experience: empathy for pain. The viewing of images of punctured or damaged body parts activates part of the same network of brain centers that are normally activated by our own sensation of pain, accounting for the feeling of physical sensation and corresponding shock upon observation of pressure or damage to the skin and limbs of others. Goya, Que hay que hacer mas? (What more is there to do?), plate 33 from Los Desastres de la Guerra (Disasters of War), etching, Biblioteque Nationale, Paris, France © Bridgeman-Giraudon/Art Resource, NY.


The implications of the discovery of mirroring mechanisms and embodied simulation for empathetic responses to images in general, and to works of visual art in particular, have not yet been assessed. Here, we address this issue and we challenge the primacy of cognition in responses to art. We propose that a crucial element of esthetic response consists of the activation of embodied mechanisms encompassing the simulation of actions, emotions and corporeal sensation, and that these mechanisms are universal. This basic level of reaction to images is essential to understanding the effectiveness both of everyday images and of works of art. Historical, cultural and other contextual factors do not preclude the importance of considering the neural processes that arise in the empathetic understanding of visual artworks.

Figure 1. Embodied simulation in esthetic experience: actions. The sense of exertion, which Michelangelo intended his figures to show as they struggle to escape from the block of stone, is effectively conveyed to the spectator. Michelangelo, Slave called Atlas, Florence, Academia (ca. 1520–1523), marble © Scala/Art Resource, NY.

Figure 2. Embodied simulation in esthetic experience: empathy for pain. The viewing of images of punctured or damaged body parts activates part of the same network of brain centers that are normally activated by our own sensation of pain, accounting for the feeling of physical sensation and corresponding shock upon observation of pressure or damage to the skin and limbs of others. Goya, Que hay que hacer mas? (What more is there to do?), plate 33 from Los Desastres de la Guerra (Disasters of War), etching, Biblioteque Nationale, Paris, France © Bridgeman-Giraudon/Art Resource, NY.

Figure 3. Embodied simulation in esthetic experience: implied gestures of the artist. The gestures that are only implicit in the marks on these works of art are corporeally felt by their spectators. (a) Jackson Pollock, Number 14: Gray (1948), enamel over gesso on paper, Yale University Gallery, The Katharine Ordway Collection © 2004 The Pollock-Krasner Foundation/Artists Rights Society (ARS), New York. (b) Lucio Fontana, Concetto Spaziale ‘Atteza’ (‘Waiting’) (1960), canvas, Tate Gallery, London © Tate Gallery, London/Art Resource, NY/ Fondazione Lucio Fontana, Milano.

Figure 4. The mirror neuron system in monkeys and humans. (a) Activation of the area F5 mirror neuron during motor-act observation. (b) Activation of the area F5 mirror neuron during action execution. For both conditions, six consecutive rasters (spike recordings) during six consecutive trials are shown. The arrows indicate the onset of observed and executed grasping. (a) and (b) modified, with permission, from Ref. [60]. (c) Somatotopy of premotor and parietal cortices as revealed by fMRI during action observation. Activation foci are projected on the lateral surface of a standard brain (MNI). Red, activation during the observation of mouth grasping; green, activation during the observation of hand grasping; blue, activation during the observation of foot kicking. Overlap of colors indicates activation foci present during observation of actions made by different effectors.
Monday, June 25, 2007
Greetings from Las Vegas and the ASSC meeting
This is the last day of the annual meeting of the Association for the Scientific Study of Consciousness. I'm going to wait and work up a few posts on some of what I think is the most interesting material over the next period of time. Las Vegas is beyond belief.... like apocalypse now.... it will take weeks for the sensory overload to fade away. Here is the Eiffel Tower in Paris, Las Vegas, and the view of the Bellagio fountains from my lunch in its restaurant (the only peaceful and quiet place in the city I found) ....




The most popular posts on MindBlog
I subscribe to a service offered at feedburner.com that lets you see details of the traffic on your blog. I am passing on the titles of the five postings that it indicates have been read the most.
Watching a computer "be nice" activates our brain's agency and goal detectors
A new desciprtion of our inner lives…
A neural network that shares a common genetic origin with human intelligence.
Social Networks - the twenty-first century science?
The pianist in the mirror - human mirror neuron systems for motor learning
Watching a computer "be nice" activates our brain's agency and goal detectors
A new desciprtion of our inner lives…
A neural network that shares a common genetic origin with human intelligence.
Social Networks - the twenty-first century science?
The pianist in the mirror - human mirror neuron systems for motor learning
New steps in memory consolidation
Paz et al show how the hippocampal-medial prefrontal cortex interactions thought to support memory consolidation are enhanced by correlated activities in regions around the hippocampus. Their introduction gives a quick summary of known steps in memory consolidation which provides context for their experiment, here are some edited clips:
The hippocampus plays a time-limited role in the formation of declarative memories, with memories gradually becoming independent of the hippocampus over time. It is believed that these remote memories are gradually transferred from the hippocampus to the neocortex for long-term storage..investigations indicate that the medial prefrontal cortex (mPFC) is critical for the consolidation of hippocampal-dependent memories. In trace-conditioning tasks for instance, hippocampal lesions cause a severe deficit when made soon after training, but not after a month, whereas mPFC lesions produce the opposite pattern of impairments. ...The role of mPFC activity in memory formation remains unclear. One possibility is that mPFC affects the transfer of hippocampal activity toward the neocortex. Consistent with this possibility, the mPFC projects to the rhinal cortices, the main route for impulse traffic into and out of the hippocampus...the present study was undertaken to test the idea that the mPFC influences memory formation by modulating interactions between the neocortex and hippocampus at the level of the rhinal cortices. To this end, we examined the relative timing of unit activity in the mPFC, PR, and ER cortices during the acquisition of a trace-conditioning task.And, here is the abstract of the paper:
Much data suggests that hippocampal–medial prefrontal cortex (mPFC) interactions support memory consolidation. This process is thought to involve the gradual transfer of transient hippocampal-dependent memories to distributed neocortical sites for long-term storage. However, hippocampal projections to the neocortex involve a multisynaptic pathway that sequentially progresses through the entorhinal and perirhinal regions before reaching the neocortex. Similarly, the mPFC influences the hippocampus via the rhinal cortices, suggesting that the rhinal cortices occupy a strategic position in this network. The present study thus tested the idea that the mPFC supports memory by facilitating the transfer of hippocampal activity to the neocortex via an enhancement of entorhinal to perirhinal communication. To this end, we simultaneously recorded mPFC, perirhinal, and entorhinal neurons during the acquisition of a trace-conditioning task in which a visual conditioned stimulus (CS) was followed by a delay period after which a liquid reward was administered. At learning onset, correlated perirhinal-entorhinal firing increased in relation to mPFC activity, but with no preferential directionality, and only after reward delivery. However, as learning progressed across days, mPFC activity gradually enhanced rhinal correlations in relation to the CS as well, and did so in a specific direction: from entorhinal to perirhinal neurons. This suggests that, at late stages of learning, mPFC activity facilitates entorhinal to perirhinal communication. Because this connection is a necessary step for the transfer of hippocampal activity to the neocortex, our results suggest that the mPFC is involved in the slow iterative process supporting the integration of hippocampal-dependent memories into neocortical networks.
Friday, June 22, 2007
Horniness of male mice - up and down with estrogen
Here is the abstract from Taziaux et al. Might this tempt pharmaceutical companies to go for a human male equivalent?
Estrogens are classically viewed as hormones that bind to intracellular receptors, which then act as transcription factors to modulate gene expression; however, they also affect many aspects of neuronal functioning by rapid nongenomic actions. Brain estrogen production can be regulated within minutes by changes in aromatase (estrogen synthase) activity as a result of calcium-dependent phosphorylations of the enzyme. To determine the effects of rapid changes in estrogen availability on male copulatory behavior, we mimicked in male mice the rapid upregulation and downregulation of brain estrogen concentration that should occur after inactivation or activation of aromatase activity. A single injection of different aromatase inhibitors [Vorozole, 1,4,6-androstatrien-3,17-dione (ATD), or its metabolite 17-OH-ATD (1,4,6-androstatrien-17ß-ol-3-one)] almost completely suppressed male sexual behavior (mounts and intromissions) expressed 10–20 min later by C57BL/6J mice but did not affect behavior in aromatase knock-out (ArKO) mice, activated by daily injections of estradiol benzoate, thereby confirming the specificity of the behavioral inhibition observed in wild-type mice. The rapid ATD-induced inhibition was reversed by the simultaneous injection of a large dose of estradiol. A single injection of estradiol to ArKO mice also activated male sexual behavior within 15 min. Thus, rapid increases or decreases in brain estrogen concentrations are followed within minutes by corresponding changes in male sexual behavior. Sexual behavior can thus be used to monitor changes in local estrogen concentrations and analyze the mechanisms mediating the rapid decline in estrogen signaling that takes place after inhibition of estrogen synthesis.
Debunking Phrenology - oscillating networks control behavior
Robert K. Night writes a perspective in Science (PDF here) on how neural networks support goal-directed behavior:


Systems neuroscience aims to understand how billions of neurons in the mammalian brain support goal-directed behavior, such as decision making. Deciphering how individual neurons respond to sensory inputs or motor decisions has focused on delineating the neural basis of these processes in discrete regions of the brain's cortex, and has provided key insights into the physiological basis of behavior. However, evidence from neuropsychological, electrophysiological, and neuroimaging studies in humans has revealed that interactions between widespread neural regions in the brain underlie fluid, organized behavior.He then summarizes work reported in three papers to:
...unravel the details of these interactions by assessing the simultaneous activity of neurons in multiple sites of the mammalian brain. The studies show that network interactions among anatomically discrete brain regions underlie cognitive processing and dispel any phrenological notion that a given innate mental faculty is based solely in just one part of the brain.
...Taken together, the three papers indicate that top-down signals between brain regions regulate the flow of information and that distributed neural networks that use oscillatory dynamics support a broad spectrum of neural processing and behavior. The results in cats and monkeys also nicely parallel findings in humans...this particular coupling mechanism is used to delineate task-specific network activity
Subscribe to:
Posts (Atom)