 (click to enlarge) Figure legend: The neurons identified by Prather and colleagues could be involved in three sensorimotor processes. a, The delayed corollary discharge of song patterns can be simultaneously compared with auditory feedback of the bird's own song, allowing tuning. b, The auditory responses (in the mirroring neurons) to songs of a neighbour might be compared with the memory of the corollary discharge produced during singing. This might allow the bird to identify an imitation by that neighbour. c, Corollary discharges while singing might be compared with a memory of the mirroring neurons' response to the parent's song. The error may then feed back to the song generator and guide vocal learning during song development, in addition to guidance from auditory input during singing (lowest arrow).
(click to enlarge) Figure legend: The neurons identified by Prather and colleagues could be involved in three sensorimotor processes. a, The delayed corollary discharge of song patterns can be simultaneously compared with auditory feedback of the bird's own song, allowing tuning. b, The auditory responses (in the mirroring neurons) to songs of a neighbour might be compared with the memory of the corollary discharge produced during singing. This might allow the bird to identify an imitation by that neighbour. c, Corollary discharges while singing might be compared with a memory of the mirroring neurons' response to the parent's song. The error may then feed back to the song generator and guide vocal learning during song development, in addition to guidance from auditory input during singing (lowest arrow).
Showing posts with label mirror neurons. Show all posts
Showing posts with label mirror neurons. Show all posts
Wednesday, January 30, 2008
Songbirds also have mirror neurons.
Mirror neurons in humans and other primates fire both when a given action is either performed or observed (see the 'mirror neuron' category in the left column of this blog.) Now Prather et al. have found similar neurons in the swamp sparrow, which like humans depends on auditory experience to learn its vocal repertoire. These forebrain neurons are specialized for auditory-vocal monitoring and have virtually the same response to a given note sequence, whether performed or heard. They also innervate striatal structures important for song learning, raising the possibility that singing-related activity in these cells is compared to auditory feedback to guide vocal learning. Here is a graphic from a New and Views article on this work by Tchernichovski1 & Wallman in the same issue of Nature.
 (click to enlarge) Figure legend: The neurons identified by Prather and colleagues could be involved in three sensorimotor processes. a, The delayed corollary discharge of song patterns can be simultaneously compared with auditory feedback of the bird's own song, allowing tuning. b, The auditory responses (in the mirroring neurons) to songs of a neighbour might be compared with the memory of the corollary discharge produced during singing. This might allow the bird to identify an imitation by that neighbour. c, Corollary discharges while singing might be compared with a memory of the mirroring neurons' response to the parent's song. The error may then feed back to the song generator and guide vocal learning during song development, in addition to guidance from auditory input during singing (lowest arrow).
(click to enlarge) Figure legend: The neurons identified by Prather and colleagues could be involved in three sensorimotor processes. a, The delayed corollary discharge of song patterns can be simultaneously compared with auditory feedback of the bird's own song, allowing tuning. b, The auditory responses (in the mirroring neurons) to songs of a neighbour might be compared with the memory of the corollary discharge produced during singing. This might allow the bird to identify an imitation by that neighbour. c, Corollary discharges while singing might be compared with a memory of the mirroring neurons' response to the parent's song. The error may then feed back to the song generator and guide vocal learning during song development, in addition to guidance from auditory input during singing (lowest arrow).
(click to enlarge) Figure legend: The neurons identified by Prather and colleagues could be involved in three sensorimotor processes. a, The delayed corollary discharge of song patterns can be simultaneously compared with auditory feedback of the bird's own song, allowing tuning. b, The auditory responses (in the mirroring neurons) to songs of a neighbour might be compared with the memory of the corollary discharge produced during singing. This might allow the bird to identify an imitation by that neighbour. c, Corollary discharges while singing might be compared with a memory of the mirroring neurons' response to the parent's song. The error may then feed back to the song generator and guide vocal learning during song development, in addition to guidance from auditory input during singing (lowest arrow).
Tuesday, January 22, 2008
The Impressionable Brain
I want to pass on this essay by Marcel Kinsbourne, which is an interesting exegesis of the implication of mirror neuron systems in our brain, especially with respect to possibly making moot the issue of brain versus environmental effects:
When the phenomenon of "mirror neurons" that fire both when a specific action is perceived and when it is intended was first reported, I was impressed by the research but skeptical about its significance...I have come to realize that mirror neurons are not only less than meets the eye but also more. Instead of being a specific specialization, they play their role as part of a fundamental design characteristic of the brain; that is, when percepts are activated, relevant intentions, memories and feelings automatically fall into place.
That an individual is likely to act in the same ways that others act is seen in the documented benefit for sports training of watching experts perform. "Emotional contagion" occurs when someone witnesses the emotional expressions of another person and therefore experiences that mood state oneself. People's viewpoints can subtly and unconsciously converge when their patterns of neural activation match, in the total absence of argument or attempts at persuasion. When people entrain with each other in gatherings, crowds, assemblies and mobs, diverse individual views reduce into a unified group viewpoint.
People's views are surreptitiously shaped by their experiences, and rationality comes limping after, downgraded to rationalization. Once opinions are established, they engender corresponding anticipations. People actively seek those experiences that corroborate their own self-serving expectations. This may be why as we grow older, we become ever more like ourselves. Insights become consolidated and biases reinforced when one only pays attention to confirming evidence. Diverse mutually contradictory "firm convictions" are the result.
If I am correct in my changed views as to what mirror neurons stand for and how representation routinely merges perception, action, memory and affect into dynamic reciprocal interaction, these views would have a bearing on currently disputed issues. Whether an effect is due to the brain or the environment would be moot if environmental causes indeed become brain causes, as the impressionable brain resonates with changing circumstances. What we experience contributes mightily to what we are and what we become.
What people experience indeed changes their brain, for better and for worse. In turn, the changed brain changes what is experienced. Regardless of its apparent stability over time, the brain is in constant flux, and constantly remodels. Heraclitus was right: "You shall not go down twice to the same river". The river will not be the same, but for that matter, neither will you. We are never the same person twice. The past is etched into the neural network, biasing what the brain is and does in the present. William Faulkner recognized this: "The past is never dead. In fact, it's not even past".

Friday, December 21, 2007
The hidden structure of over-imitation
Human children, unlike chimpanzees, will copy unnecessary or arbitrary parts of an action sequence they observe in adults, Lyons et al. term this process overimitation and suggest in an open access article with the title of this post that it reveals a hidden structure behind how children learn to attribute causality. Here is their abstract, and a graphic showing one of the three puzzle boxes used in the experiements:
Young children are surprisingly judicious imitators, but there are also times when their reproduction of others' actions appears strikingly illogical. For example, children who observe an adult inefficiently operating a novel object frequently engage in what we term overimitation, persistently reproducing the adult's unnecessary actions. Although children readily overimitate irrelevant actions that even chimpanzees ignore, this curious effect has previously attracted little interest; it has been assumed that children overimitate not for theoretically significant reasons, but rather as a purely social exercise. In this paper, however, we challenge this view, presenting evidence that overimitation reflects a more fundamental cognitive process.We show that children who observe an adult intentionally manipulating a novel object have a strong tendency to encode all of the adult's actions as causally meaningful, implicitly revising their causal understanding of the object accordingly. This automatic causal encoding process allows children to rapidly calibrate their causal beliefs about even the most opaque physical systems, but it also carries a cost. When some of the adult's purposeful actions are unnecessary—even transparently so—children are highly prone to mis-encoding them as causally significant. The resulting distortions in children's causal beliefs are the true cause of overimitation, a fact that makes the effect remarkably resistant to extinction. Despite countervailing task demands, time pressure, and even direct warnings, children are frequently unable to avoid reproducing the adult's irrelevant actions because they have already incorporated them into their representation of the target object's causal structure.

Tuesday, November 20, 2007
Impairment of action chains in autism.
When we observe the start of an action sequence that can end in two possible ways (in the figure shown a piece of food is placed in the mouth or in a container on the shoulder) appropriate sympathetic muscle EMG signals are detected at the start of the sequence. Thus, if the sequence will end in food to the mouth, activity is observed in the mouth-opening mylohyoid (MH) muscle at the onset. Rizzolatti and collaborators find that typically developing children show an activation of their MH muscle already when they observe the experimenter's initial motor act, food reaching. This activation reflects their understanding of the final goal of the observed action. In children with autism this action-understanding motor activation is lacking. Further, when typically developing children actually perform the observed action, MH muscle activation is observed at the very beginning of the sequence, while in children with autism, the activation is not observed until immediately before the muscle is actually used.
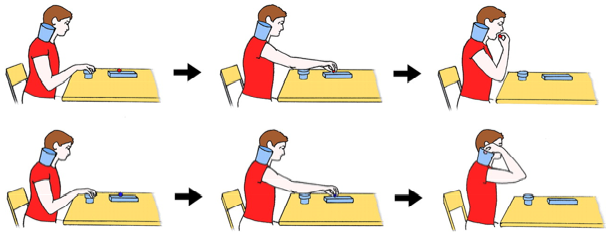 Figure - Schematic representation of the tasks. (Upper) The individual reaches for a piece of food located on a touch-sensitive plate, grasps it, brings it to the mouth, and finally eats it. (Lower) The individual reaches for a piece of a paper located on the same plate, grasps it, and puts into a container placed on the shoulder.
Figure - Schematic representation of the tasks. (Upper) The individual reaches for a piece of food located on a touch-sensitive plate, grasps it, brings it to the mouth, and finally eats it. (Lower) The individual reaches for a piece of a paper located on the same plate, grasps it, and puts into a container placed on the shoulder.
They suggest that high-functioning autistic children may understand the intentions of others cognitively but lack the mechanism for understanding them experientially because they lack the chains of action-constrained neurons that code specific motor acts (e.g., grasping) according to the final goal of the action in which the motor act is embedded.
 Figure - Schematic representation of the tasks. (Upper) The individual reaches for a piece of food located on a touch-sensitive plate, grasps it, brings it to the mouth, and finally eats it. (Lower) The individual reaches for a piece of a paper located on the same plate, grasps it, and puts into a container placed on the shoulder.
Figure - Schematic representation of the tasks. (Upper) The individual reaches for a piece of food located on a touch-sensitive plate, grasps it, brings it to the mouth, and finally eats it. (Lower) The individual reaches for a piece of a paper located on the same plate, grasps it, and puts into a container placed on the shoulder.They suggest that high-functioning autistic children may understand the intentions of others cognitively but lack the mechanism for understanding them experientially because they lack the chains of action-constrained neurons that code specific motor acts (e.g., grasping) according to the final goal of the action in which the motor act is embedded.
Monday, October 08, 2007
Plasticity and learning in the human mirror neuron system
I pass on a review by Welberg of an interesting study by Catmur et al. [Catmur, C., Walsh, V. & Heyes, C. Sensorimotor learning configures the human mirror system. Curr. Biol. 17, 1527–1531 (2007)]:
Neurons in the frontoparietal mirror system fire when one performs an action and when one observes someone else performing that same action. This system is thought to have a role in social cognition and, perhaps, in language acquisition. How the mirror neurons map sensory input onto its motor representation is unknown, but Catmur et al. demonstrate that these representations are not innate and can be altered by training.
The authors used transcranial magnetic stimulation (TMS) to stimulate the motor cortex of volunteers who were watching a video of a hand. When the volunteers watched the hand's index finger move, the TMS-induced motor-evoked potential (MEP) was greater in the abductor muscle of their own index finger than when they watched the little finger move; conversely, the MEP of their little finger's abductor muscle was greatest when they watched the little finger move. In other words, a muscle showed MEP enhancement when its owner watched a movement that is normally performed by that muscle; this 'mirror effect' is thought to reflect activity of the mirror neuron system.
Half of the volunteers then underwent incongruent training trials, in which they were asked to extend their little finger if the video showed a hand extending the index finger, and vice versa. People in congruent trials simply had to repeat the movement they saw in the video. The incongruent trials were assumed to train the mirror system to associate an observed finger movement with movement of a different finger of the volunteer's own hand.
Measuring TMS-induced MEPs after training, the authors found that volunteers who had undergone the incongruent training now showed greater MEPs in the muscle of one finger when watching the 'wrong' finger move in the video, indicating that a reversal of muscle-specific MEP enhancement during action observation had taken place.
This study shows that the 'mirror properties' of the mirror system are not innate. Rather, they can be trained, through sensorimotor experience, to transform observation into action. These findings imply that insufficient social interaction and consequent inadequate sensory experience might affect the development of the mirror neuron system, for example, in children with autism.
Tuesday, July 10, 2007
When watching a tactile stroke is the same as receiving one.
An interesting report from Banissy and Ward (PDF here)... People who have 'mirror-touch' synesthesia, when watching another person being touched, have the same experience as being touched themselves...an extreme form of empathy! They developed a protocol to provide evidence for the authenticity of this form of synesthesia, in which participants have difficulty in discriminating between actual and synesthetic touch. Their abstract:
Watching another person being touched activates a similar neural circuit to actual touch and, for some people with 'mirror-touch' synesthesia, can produce a felt tactile sensation on their own body. In this study, we provide evidence for the existence of this type of synesthesia and show that it correlates with heightened empathic ability. This is consistent with the notion that we empathize with others through a process of simulation.
Thursday, July 05, 2007
Where the brain understands animate agents..
Wheatley et al offer an interesting study in a recent issue of Psychological Science (vol 18, pg 469, 2007, PDF here). Here is the abstract and two figures:
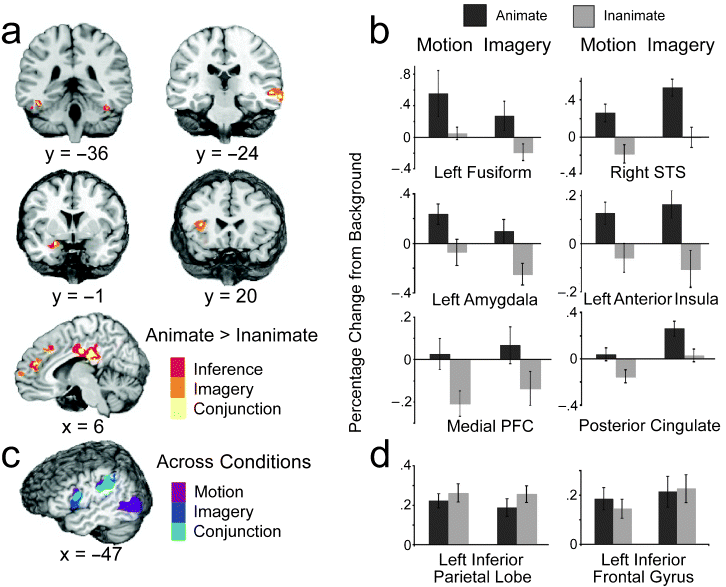
How people understand the actions of animate agents has been vigorously debated. This debate has centered on two hypotheses focused on anatomically distinct neural substrates: The mirror-system hypothesis proposes that the understanding of others is achieved via action simulation, and the social-network hypothesis proposes that such understanding is achieved via the integration of critical biological properties (e.g., faces, affect). In this study, we assessed the areas of the brain that were engaged when people interpreted and imagined moving shapes as animate or inanimate. Although observing and imagining the moving shapes engaged the mirror system, only activation of the social network was modulated by animacy.Lateral and medial views of the social network (top, highlighted in yellow) and mirror system (bottom, highlighted in blue). The social network includes areas associated with biological motion (superior temporal sulcus, labeled "1"), biological form (lateral fusiform gyrus, labeled "6"), mentalizing (medial prefrontal cortex and posterior cingulate, labeled "3" and "4," respectively), and affective processing (insula and amygdala, labeled "2" and "5," respectively). The mirror system consists of the inferior parietal cortex (labeled "7") and the ventral-premotor/inferior-frontal cortex (labeled "8").
Experimental results. The brain slices in (a) depict areas of the social network that were more active when moving shapes were inferred (red) or imagined (orange) as animate than when they were inferred or imagined as inanimate. Yellow areas were more active for both animate inference and imagery ("conjunction"). The graph in (b) displays the average hemodynamic responses within the conjunction areas as a function of animacy (animate, inanimate) and condition (motion, imagery). (Results are not shown for the posterior insula, although this was also a conjunction area.) The illustration in (c) shows areas of the mirror system that were more active when subjects watched and made inferences about the moving shapes (purple) and when they imagined (dark blue) the moving shapes relative to when they viewed the backgrounds alone; light-blue areas were more active during both the motion and imagery conditions ("conjunction") than in the background condition. The graph in (d) shows the average hemodynamic responses of the conjunction mirror areas as a function of animacy and condition. For purposes of illustration, all group data are presented on the N27 (AFNI software) brain. Error bars represent standard errors. STS = superior temporal sulcus; PFC = prefrontal cortex.

Tuesday, June 26, 2007
Esthetic experience and mirror neurons - motion, emotion and empathy
David Freedberg (Art History, Columbia Univ.) and Vittorio Gallese (Neuroscience, Parma) offer an interesting perspective on possible relationships between mirror neuron systems and artistic experience. Here is their abstract (and PDF of article is here.), followed by some of their illustrations.

Figure 2. Embodied simulation in esthetic experience: empathy for pain. The viewing of images of punctured or damaged body parts activates part of the same network of brain centers that are normally activated by our own sensation of pain, accounting for the feeling of physical sensation and corresponding shock upon observation of pressure or damage to the skin and limbs of others. Goya, Que hay que hacer mas? (What more is there to do?), plate 33 from Los Desastres de la Guerra (Disasters of War), etching, Biblioteque Nationale, Paris, France © Bridgeman-Giraudon/Art Resource, NY.


The implications of the discovery of mirroring mechanisms and embodied simulation for empathetic responses to images in general, and to works of visual art in particular, have not yet been assessed. Here, we address this issue and we challenge the primacy of cognition in responses to art. We propose that a crucial element of esthetic response consists of the activation of embodied mechanisms encompassing the simulation of actions, emotions and corporeal sensation, and that these mechanisms are universal. This basic level of reaction to images is essential to understanding the effectiveness both of everyday images and of works of art. Historical, cultural and other contextual factors do not preclude the importance of considering the neural processes that arise in the empathetic understanding of visual artworks.

Figure 1. Embodied simulation in esthetic experience: actions. The sense of exertion, which Michelangelo intended his figures to show as they struggle to escape from the block of stone, is effectively conveyed to the spectator. Michelangelo, Slave called Atlas, Florence, Academia (ca. 1520–1523), marble © Scala/Art Resource, NY.

Figure 2. Embodied simulation in esthetic experience: empathy for pain. The viewing of images of punctured or damaged body parts activates part of the same network of brain centers that are normally activated by our own sensation of pain, accounting for the feeling of physical sensation and corresponding shock upon observation of pressure or damage to the skin and limbs of others. Goya, Que hay que hacer mas? (What more is there to do?), plate 33 from Los Desastres de la Guerra (Disasters of War), etching, Biblioteque Nationale, Paris, France © Bridgeman-Giraudon/Art Resource, NY.

Figure 3. Embodied simulation in esthetic experience: implied gestures of the artist. The gestures that are only implicit in the marks on these works of art are corporeally felt by their spectators. (a) Jackson Pollock, Number 14: Gray (1948), enamel over gesso on paper, Yale University Gallery, The Katharine Ordway Collection © 2004 The Pollock-Krasner Foundation/Artists Rights Society (ARS), New York. (b) Lucio Fontana, Concetto Spaziale ‘Atteza’ (‘Waiting’) (1960), canvas, Tate Gallery, London © Tate Gallery, London/Art Resource, NY/ Fondazione Lucio Fontana, Milano.

Figure 4. The mirror neuron system in monkeys and humans. (a) Activation of the area F5 mirror neuron during motor-act observation. (b) Activation of the area F5 mirror neuron during action execution. For both conditions, six consecutive rasters (spike recordings) during six consecutive trials are shown. The arrows indicate the onset of observed and executed grasping. (a) and (b) modified, with permission, from Ref. [60]. (c) Somatotopy of premotor and parietal cortices as revealed by fMRI during action observation. Activation foci are projected on the lateral surface of a standard brain (MNI). Red, activation during the observation of mouth grasping; green, activation during the observation of hand grasping; blue, activation during the observation of foot kicking. Overlap of colors indicates activation foci present during observation of actions made by different effectors.
Thursday, June 14, 2007
More on the mirror "hearing-doing" system
Here is a more extended commentary on a paper I mentioned in my Jan. 30 post "The pianist in the mirror.." D'Ausilio (PDF here) discusses this work of Lahav et al. showing an auditory mirror area in the left inferior frontal gyrus for complex and newly acquired actions. In addition to rote auditory-motor mapping for learning and online execution control, this mirror mechanism might subserve other evolutionary critical functions like action recognition, and interindividual emotional resonance. This auditory mirror-like property seems indeed to be valid for a wide range of functions that in turn elicit very different behaviors.
Tuesday, May 29, 2007
Neural correlates of social cognition
Vittorio Gallese writes an interesting summary in Proc. Roy. Soc. B titled "Before and below ‘theory of mind’: embodied simulation and the neural correlates of social cognition." (PDF here). His abstract:
The automatic translation of folk psychology into newly formed brain modules specifically dedicated to mind-reading and other social cognitive abilities should be carefully scrutinized. Searching for the brain location of intentions, beliefs and desires—as such—might not be the best epistemic strategy to disclose what social cognition really is. The results of neurocognitive research suggest that in the brain of primates, mirror neurons, and more generally the premotor system, play a major role in several aspects of social cognition, from action and intention understanding to language processing. This evidence is presented and discussed within the theoretical frame of an embodied simulation account of social cognition. Embodied simulation and the mirror neuron system underpinning it provide the means to share communicative intentions, meaning and reference, thus granting the parity requirements of social communication.
Friday, May 11, 2007
Integrating different theory of mind models.
Keysers and Gazzola propose a speculative model (Trends in Cognitive Sciences
Volume 11, Issue 5, May 2007, Pages 194-196. PDF here) that attempts to integrate the perspective of two polarized camps:
Volume 11, Issue 5, May 2007, Pages 194-196. PDF here) that attempts to integrate the perspective of two polarized camps:
The simulation camp focuses on so-called shared circuits (SCs) that are involved in one's own actions, sensations and emotions and in perceiving those of others. The theory of mind (ToM) camp emphasizes the role of midline structures in mentalizing about the states of others.
Social cognitions range from the intuitive examples studied by simulationists to the reflective ones used by ToM investigators. Witnessing someone drink a glass of milk with a face contracting in an expression of disgust is an example at the intuitive extreme of this continuum. In such cases, premotor and parietal areas for actions, the insula for emotions and and SII for sensations form SCs that translate the bodily states of others into the neural language of our own states. These SCs seem to implement a pre-reflective, intuitive and empathic level of representation: neural activity in these areas does not require specific instructions that encourage conscious reflections.
Thinking about what gift would please a foreign colleague is an example at the more reflective extreme. In such cases, we must browse consciously through what we know about his country and culture to deduce what he might like. Such explicit knowledge about the inner life of others is the product of reflecting upon the states of others and is linked with activity in midline structures and the temporoparietal junction. False beliefs are prototypical examples of such reflective representations.
They suggest a working hypothesis:
While dealing with states of the self, areas of the SCs represent pre-reflective bodily states. If asked to introspect and report these states, subjects additionally activate (v)mPFC structures. When dealing with states of other individuals, activity in SCs might represent the empathic transformation of the bodily states of others into pre-reflective neural representations of similar states of the self. These simulated pre-reflective representations correlate with empathy and might provide an intuitive understanding of what goes on in others. If asked to reflect on the states of others, the pathways that are normally used to reflect on the bodily representations of the self are now used on simulated bodily states of others, leading to simulated reflective representations. Thus, SCs and midline structures form an integrated system that applies to cases where we perceive the other as similar enough for simulation to be useful. In this view, both SCs and vmPFC reflect simulation, albeit at different levels (pre-reflective versus reflective), rather than radically different processes (SC versus ToM).Illustration of the model (click to enlarge). The self is shown in red, the other is shown in green and candidate brain areas that are thought to implement representation are shown in blue. During our own experiences, pre-reflective representations can lead, through introspection, to reflective representations (red). While witnessing the states of others, mirroring leads to activations that simulate pre-reflective representations of our own bodily states. A process of social introspection, utilizing the mechanisms of introspection, activates representations that simulate reflective representations of our own bodily states. A more cognitive route leads to more abstract knowledge about the other that escapes from the constraints of our own experiences.

Monday, April 30, 2007
The Myth of Mirror Neurons?
In an article in a special issue of Slate devoted to the brain (well worth checking over...I'll give some links to articles in the Slate issue in subsequent posts), Gopnik argues that excitement over the discovery of mirror neurons in our brains (the subject of a number of blog posts and my lecture posted earlier...) is generating a new scientific myth. Like a traditional myth, it captures intuitions about the human condition through vivid metaphors. Some clips:
It didn't take long for scientists and science writers to speculate that mirror neurons might serve as the physiological basis for a wide range of social behaviors, from altruism to art appreciation. Headlines like "Cells That Read Minds" or "How Brain's 'Mirrors' Aid Our Social Understanding" tapped into our intuitions about connectedness. Maybe this cell, with its mellifluous name, gives us our special capacity to understand one another—to care, to learn, and to communicate. Could mirror neurons be responsible for human language, culture, empathy, and morality?.The evidence for individual mirror neurons comes entirely from studies of macaque monkeys. That's because you can't find these cells without inserting electrodes directly (though painlessly) into individual neurons in the brains of living animals. These studies haven't been done with chimpanzees, let alone humans.
The trouble is that macaque monkeys don't have language, they don't have culture, and they don't understand other animals' minds. In fact, careful experiments show that they don't even systematically imitate the actions of other monkeys—and they certainly don't imitate in the prolific way that the youngest human children do. Even chimpanzees, who are much more cognitively sophisticated than macaques, show only very limited abilities in these areas. The fact that macaques have mirror neurons means that these cells can't by themselves explain our social behavior.

Friday, April 06, 2007
Motor cortex for the hand and numerical counting
The April 2007 issue of the Journal of Cognitive Neuroscience has interesting articles on this topic by Andres et al. and Sato et al. Their abstracts:
Andres et al.
Sato et al.
Andres et al.
The finding that number processing activates a cortical network partly overlapping that recruited for hand movements has renewed interest in the relationship between number and finger representations. Further evidence about a possible link between fingers and numbers comes from developmental studies showing that finger movements play a crucial role in learning counting. However, increased activity in hand motor circuits during counting may unveil unspecific processes, such as shifting attention, reciting number names, or matching items with a number name. To address this issue, we used transcranial magnetic stimulation to measure changes in corticospinal (CS) excitability during a counting task performed silently and using either numbers or letters of the alphabet to enumerate items. We found an increased CS excitability of hand muscles during the counting task, irrespective of the use of numbers or letters, whereas it was unchanged in arm and foot muscles. Control tasks allowed us to rule out a possible influence of attention allocation or covert speech on CS excitability increase of hand muscles during counting. The present results support a specific involvement of hand motor circuits in counting because no CS changes were found in arm and foot muscles during the same task. However, the contribution of hand motor areas is not exclusively related to number processing because an increase in CS excitability was also found when letters were used to enumerate items. This finding suggests that hand motor circuits are involved whenever items have to be put in correspondence with the elements of any ordered series.
Sato et al.
Developmental and cross-cultural studies show that finger counting represents one of the basic number learning strategies. However, despite the ubiquity of such an embodied strategy, the issue of whether there is a neural link between numbers and fingers in adult, literate individuals remains debated. Here, we used transcranial magnetic stimulation to study changes of excitability of hand muscles of individuals performing a visual parity judgment task, a task not requiring counting, on Arabic numerals from 1 to 9. Although no modulation was observed for the left hand muscles, an increase in amplitude of motor-evoked potentials was found for the right hand muscles. This increase was specific for smaller numbers (1 to 4) as compared to larger numbers (6 to 9). These findings indicate a close relationship between hand/finger and numerical representations.
Thursday, April 05, 2007
Self and social cognition in the brain
Uddin et al. speculate in Trends in Cognitive Science (Volume 11, Issue 4 , April 2007, Pages 153-157) about the respective roles of cortical midline structures (CMS) and frontoparietal mirror neurons (MNS) in evaluation of self and others. Below I relay one of their summary paragraphs, and the PDF which contains images of relevant brain regions can be obtained HERE.
It has recently been proposed that internally oriented processes that focus on one's own or others’ mental states rely on cortical midline structures, whereas externally focused processes based on one's own or others’ visible features and actions rely on lateral frontoparietal networks. We suggest here a similar distinction, which might further reconcile disparate findings with regard to the various proposed functions of cortical midline structures, while incorporating what is known about the role of the human mirror-neuron system in social cognition. Whereas there is mounting evidence that the right frontoparietal system is involved in representing the physical, embodied self (in addition to its role in understanding the actions of others), the cortical midline structures that comprise the default-mode network seem to be more involved in maintaining a self-representation in evaluative terms, which requires self-referential processing and understanding of others’ mental states. We speculate that the CMS might support evaluative simulation in the same way that the MNS supports motor simulation. This distinction serves as a practical division of labor between two networks that are specialized for two related processes that are crucial to navigating the social world. The mirror-neuron system provides the essential physical other-to-self mapping that is necessary for comprehending physical actions of intentional agents, whereas cortical midline structures maintain and support processes that are related to understanding complex psychological aspects of others, such as attitudes, perhaps by simulation of one's own attitudes.
Tuesday, April 03, 2007
Mirror Neurons - gee whiz popular video
A engaging and easy to watch popularized version of material I've written about earlier (see 1/20/07; 12/19/06; and my lecture 10/27/06)).
Tuesday, February 27, 2007
Neuroscience will change society
Interesting reflections by Marco Iacoboni, one of the discoverers of mirror neurons, which have been mentioned a number of times in this blog:
"...a concept that emerges from recent neuroscience research is that humans are "wired for empathy". We have cells in our brains that make us understand each other in a simple, unmediated, automatic manner. But, if our neurobiology makes us wired for empathy, why is our world so full of atrocities?
The explanation for this apparent paradox is probably as follows. The neurobiological mechanisms that make us wired for empathy work at a pre-reflective, automatic, implicit level. Our societies are built on deliberate, reflective, explicit discourse. The two different levels of implicit and explicit mental processes rarely intersect; indeed there is evidence that they can often dissociate. This is probably why the massive belief systems—from religious to political ones—that operate at the deliberate, reflective level are able to divide us in such a powerful way even though our neurobiology should bring us together.
The good news is that the awareness of neurobiological mechanisms that make us wired for empathy is entering the public discourse.... This awareness won't go away and will seep through the reflective level of our mental processes. Indeed, people seem to have an intuitive understanding of how neural mechanisms for empathy work. It seems that people 'recognize' how their brain works, when they are told about it. People can finally articulate what they already 'knew' at a pre-reflective level. My optimism is that this explicit level of understanding of our empathic nature will at some point dissolve the massive belief systems that dominate our societies and that threaten to destroy us."
"...a concept that emerges from recent neuroscience research is that humans are "wired for empathy". We have cells in our brains that make us understand each other in a simple, unmediated, automatic manner. But, if our neurobiology makes us wired for empathy, why is our world so full of atrocities?
The explanation for this apparent paradox is probably as follows. The neurobiological mechanisms that make us wired for empathy work at a pre-reflective, automatic, implicit level. Our societies are built on deliberate, reflective, explicit discourse. The two different levels of implicit and explicit mental processes rarely intersect; indeed there is evidence that they can often dissociate. This is probably why the massive belief systems—from religious to political ones—that operate at the deliberate, reflective level are able to divide us in such a powerful way even though our neurobiology should bring us together.
The good news is that the awareness of neurobiological mechanisms that make us wired for empathy is entering the public discourse.... This awareness won't go away and will seep through the reflective level of our mental processes. Indeed, people seem to have an intuitive understanding of how neural mechanisms for empathy work. It seems that people 'recognize' how their brain works, when they are told about it. People can finally articulate what they already 'knew' at a pre-reflective level. My optimism is that this explicit level of understanding of our empathic nature will at some point dissolve the massive belief systems that dominate our societies and that threaten to destroy us."
Tuesday, January 30, 2007
The pianist in the mirror - human mirror neuron systems for motor learning
A Society for Neuroscience news release (pointed out to me by colleague Robin Chapman) focuses on a new imaging study that shows that when we learn a new action with associated sounds, the brain quickly makes links between regions responsible for performing the action and those associated with the sound. The findings may contribute to understanding how we acquire language and how we think of actions if we only hear their sounds. Lahav et al. taught nine subjects with no previous musical training to play a five-note, 24-second song on a keyboard. Then they ran functional magnetic resonance imaging (fMRI) scans while the subjects listened to the song they had just learned, a different song using the same five notes, and a third song made up of additional notes. From Lahav et al.'s abstract: "Although subjects listened to the music without performing any movements, activation was found bilaterally in the frontoparietal motor-related network (including Broca's area, the premotor region, the intraparietal sulcus, and the inferior parietal region), consistent with neural circuits that have been associated with action observations, and may constitute the human mirror neuron system. Presentation of the practiced notes in a different order activated the network to a much lesser degree, whereas listening to an equally familiar but motorically unknown music did not activate this network. These findings support the hypothesis of a "hearing–doing" system that is highly dependent on the individual's motor repertoire, gets established rapidly, and consists of Broca's area as its hub."

Figure 1. Action–listening illustration. A, Music performance can be viewed as a complex sequence of both actions and sounds, in which sounds are made by actions. B, The sound of music one knows how to play can be reflected, as if in a mirror, in the corresponding motor representations.

Figure 2. Action–listening activation. A, B, Extensive bilateral activation in the frontoparietal motor-related brain regions was observed when subjects listened to the trained-music they knew how to play (A), but not when they listened to the never-learned untrained-different-notes-music (B). C, Activation maps are shown in areas that were significantly more active during listening to the trained-music versus the untrained-different-notes-music. L, Left; R, right.

Figure 1. Action–listening illustration. A, Music performance can be viewed as a complex sequence of both actions and sounds, in which sounds are made by actions. B, The sound of music one knows how to play can be reflected, as if in a mirror, in the corresponding motor representations.
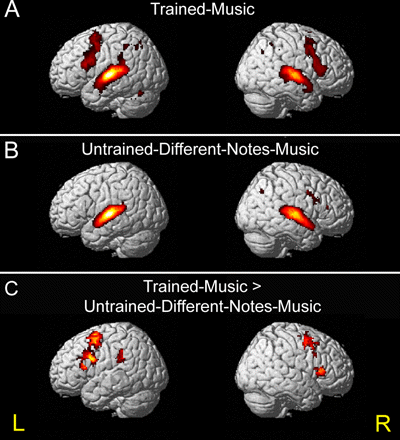
Figure 2. Action–listening activation. A, B, Extensive bilateral activation in the frontoparietal motor-related brain regions was observed when subjects listened to the trained-music they knew how to play (A), but not when they listened to the never-learned untrained-different-notes-music (B). C, Activation maps are shown in areas that were significantly more active during listening to the trained-music versus the untrained-different-notes-music. L, Left; R, right.
Tuesday, December 19, 2006
An auditory-motor 'mirror" system is engaged by positive emotions.
More on mirror systems from Warren et al in Journal of Neuroscience. Edited clips from their paper:
Social interaction relies on the ability to react to communication signals. Although cortical sensory–motor "mirror" networks are thought to play a key role in visual aspects of primate communication, evidence for a similar generic role for auditory–motor interaction in primate nonverbal communication is lacking.
In this functional magnetic resonance imaging (fMRI) study, the authors investigated cortical regions responsive to both the perception of human vocalizations and the voluntary generation of facial expressions. In four auditory–perceptual conditions, subjects listened passively, without overt motor response, to nonverbal emotional vocalizations conveying two positive-valence emotions, amusement and triumph, and two negative-valence emotions, fear and disgust. Use of nonverbal, rather than verbal, vocalizations optimized recognizability of emotional content and avoided confounds of phonological and verbal content. In a facial movement condition, subjects performed voluntary smiling movements in the absence of auditory input. They hypothesized that cortical regions showing combined auditory–perceptual and motor responses would be located within premotor and motor cortical regions.

Figure legend: Brain regions demonstrating auditory–motor mirror responses. A shows regions (red) displaying a significant modulatory effect of emotion category on perceptual activation. B shows regions (light green) displaying significant activation during voluntary facial movements (motor > baseline). C, A masked inclusively in B shows regions (dark green) displaying both a significant modulatory effect of emotion category on perceptual activation and significant activation during voluntary facial movements.

Figure legend: Correlations with emotional valence and arousal in brain regions demonstrating auditory–motor mirror responses. Left, Regions (green) displaying both a significant modulatory effect of emotion category on perceptual activation and significant activation during voluntary facial movements as shown in the figure above. Right, Regions demonstrating a significant positive correlation between hemodynamic responses and emotional valence (red), emotional arousal (blue), or both (purple).
The authors demonstrated that a network of human premotor cortical regions activated during facial movement is also involved in auditory processing of affective nonverbal vocalizations. Within this auditory–motor mirror network, distinct functional subsystems respond preferentially to emotional valence and arousal properties of heard vocalizations. Positive emotional valence enhanced activation in a left posterior inferior frontal region involved in representation of prototypic actions, whereas increasing arousal enhanced activation in presupplementary motor area cortex involved in higher-order motor control. Their findings demonstrate that listening to nonverbal vocalizations can automatically engage preparation of responsive orofacial gestures, an effect that is greatest for positive-valence and high-arousal emotions. The automatic engagement of responsive orofacial gestures by emotional vocalizations suggests that auditory–motor interactions provide a fundamental mechanism for mirroring the emotional states of others during primate social behavior.
Motor facilitation by positive vocal emotions suggests a basic neural mechanism for establishing cohesive bonds within primate social groups.
Social interaction relies on the ability to react to communication signals. Although cortical sensory–motor "mirror" networks are thought to play a key role in visual aspects of primate communication, evidence for a similar generic role for auditory–motor interaction in primate nonverbal communication is lacking.
In this functional magnetic resonance imaging (fMRI) study, the authors investigated cortical regions responsive to both the perception of human vocalizations and the voluntary generation of facial expressions. In four auditory–perceptual conditions, subjects listened passively, without overt motor response, to nonverbal emotional vocalizations conveying two positive-valence emotions, amusement and triumph, and two negative-valence emotions, fear and disgust. Use of nonverbal, rather than verbal, vocalizations optimized recognizability of emotional content and avoided confounds of phonological and verbal content. In a facial movement condition, subjects performed voluntary smiling movements in the absence of auditory input. They hypothesized that cortical regions showing combined auditory–perceptual and motor responses would be located within premotor and motor cortical regions.

Figure legend: Brain regions demonstrating auditory–motor mirror responses. A shows regions (red) displaying a significant modulatory effect of emotion category on perceptual activation. B shows regions (light green) displaying significant activation during voluntary facial movements (motor > baseline). C, A masked inclusively in B shows regions (dark green) displaying both a significant modulatory effect of emotion category on perceptual activation and significant activation during voluntary facial movements.

Figure legend: Correlations with emotional valence and arousal in brain regions demonstrating auditory–motor mirror responses. Left, Regions (green) displaying both a significant modulatory effect of emotion category on perceptual activation and significant activation during voluntary facial movements as shown in the figure above. Right, Regions demonstrating a significant positive correlation between hemodynamic responses and emotional valence (red), emotional arousal (blue), or both (purple).
The authors demonstrated that a network of human premotor cortical regions activated during facial movement is also involved in auditory processing of affective nonverbal vocalizations. Within this auditory–motor mirror network, distinct functional subsystems respond preferentially to emotional valence and arousal properties of heard vocalizations. Positive emotional valence enhanced activation in a left posterior inferior frontal region involved in representation of prototypic actions, whereas increasing arousal enhanced activation in presupplementary motor area cortex involved in higher-order motor control. Their findings demonstrate that listening to nonverbal vocalizations can automatically engage preparation of responsive orofacial gestures, an effect that is greatest for positive-valence and high-arousal emotions. The automatic engagement of responsive orofacial gestures by emotional vocalizations suggests that auditory–motor interactions provide a fundamental mechanism for mirroring the emotional states of others during primate social behavior.
Motor facilitation by positive vocal emotions suggests a basic neural mechanism for establishing cohesive bonds within primate social groups.
Thursday, November 02, 2006
Self recognition - humans, apes, dolphins...and now elephants
Plotnik, de Waal, and Reiss report in PNAS that mirror self recognition (MSR), considered an indicator of self awareness, can be observed in Asian elephants. Slightly edited from their abstract: "MSR is thought to correlate with higher forms of empathy and altruistic behavior. Apart from humans and apes, dolphins and elephants are also known for such capacities. After the recent discovery of MSR in dolphins (Tursiops truncatus), elephants thus were the next logical candidate species. The authors exposed three Asian elephants (Elephas maximus) to a large mirror to investigate their responses. Animals that possess MSR typically progress through four stages of behavior when facing a mirror: (i) social responses, (ii) physical inspection (e.g., looking behind the mirror), (iii) repetitive mirror-testing behavior, and (iv) realization of seeing themselves. Visible marks and invisible sham-marks were applied to the elephants' heads to test whether they would pass the litmus "mark test" for MSR in which an individual spontaneously uses a mirror to touch an otherwise imperceptible mark on its own body. Here, we report a successful MSR elephant study and report striking parallels in the progression of responses to mirrors among apes, dolphins, and elephants. These parallels suggest convergent cognitive evolution most likely related to complex sociality and cooperation.

Above video (click the rectangle to play) was taken from the Elmo lipstick camera embedded in the mirror on Happy's first day of marking. Happy repetitively touches the mark or the area immediately around the mark with her trunk while in full view of the mirror. She never touches the sham-mark during this session.
Above video (click the rectangle to play) was taken from the Elmo lipstick camera embedded in the mirror on Happy's first day of marking. Happy repetitively touches the mark or the area immediately around the mark with her trunk while in full view of the mirror. She never touches the sham-mark during this session.
Friday, October 27, 2006
Mirroring Minds (A talk I gave at Univ. Wisc. on 10/24)
An eclectic group of people gathers on Tuesday noon every week at the "Chaos and Complexity" seminar organized by Clint Sprott of the University of Wisconsin Madison Physics Department and Robin Chapman, a developmental psychologist and poet. They asked me to give a talk for that group and this is a link to a minimally edited version: Mirroring Minds: Are Mirror Neuron Systems the DNA of Psychology or a Red Herring? The web essays on my wesite "The I Illusion" and "The Beast Within" are previous talks I have given to this group. I had hoped to get as satisfying a bottom line on the "social brain" as I felt was obtained in those earlier talks, but it didn't really happen. Still, people were very positive about the talk and the discussion was excellent, so I decided to give you the above link to it.
Subscribe to:
Posts (Atom)