Attractive faces can be considered to be a form of visual reward. Previous imaging studies have reported activity in reward structures including orbitofrontal cortex and nucleus accumbens during presentation of attractive faces. Given that these stimuli appear to act as rewards, we set out to explore whether it was possible to establish conditioning in human subjects by pairing presentation of arbitrary affectively neutral stimuli with subsequent presentation of attractive and unattractive faces. Furthermore, we scanned human subjects with functional magnetic resonance imaging (fMRI) while they underwent this conditioning procedure to determine whether a reward-prediction error signal is engaged during learning with attractive faces as is known to be the case for learning with other types of reward such as juice and money. Subjects showed changes in behavioral ratings to the conditioned stimuli (CS) when comparing post- to preconditioning evaluations, notably for those CSs paired with attractive female faces. We used a simple Rescorla-Wagner learning model to generate a reward-prediction error signal and entered this into a regression analysis with the fMRI data. We found significant prediction error-related activity in the ventral striatum during conditioning with attractive compared with unattractive faces. These findings suggest that an arbitrary stimulus can acquire conditioned value by being paired with pleasant visual stimuli just as with other types of reward such as money or juice. This learning process elicits a reward-prediction error signal in a main target structure of dopamine neurons: the ventral striatum. The findings we describe here may provide insights into the neural mechanisms tapped into by advertisers seeking to influence behavioral preferences by repeatedly exposing consumers to simple associations between products and rewarding visual stimuli such as pretty faces.Legend - Prediction error related activity in the nucleus accumbens. A: voxels in the nucleus accumbens were significantly activated in a contrast of prediction error signals for attractive faces vs. unattractive faces, voxels in yellow are significant at P <>
Thursday, April 12, 2007
How pretty faces sell boring products - conditioning our brain's reward system
Bray and O'Doherty's report in the Journal of Neurophysiology illustrates the biology behind the selling power of beautiful models in advertisements for mundane products, and should be of interest to neuro-marketers everywhere. PDF here. Their abstract:

MindBlog back in Madison, Wisconsin

Turns out I got back just in time for what is hopefully the last snowstorm of the winter. This reminds me why I was in Ft. Lauderdale for the past five months.....

Wednesday, April 11, 2007
"Nest" cells in the mouse brain. Category knowledge shown.
An interesting article in PNAS from Lin et al. (PDF here). Their abstract:
As important as memory is to our daily functions, the ability to extract fundamental features and commonalities from various episodic experiences and to then generalize them into abstract concepts is even more crucial for both humans and animals to adapt to novel and complex situations. Here, we report the neural correlates of the abstract concept of nests or beds in mice. Specifically, we find hippocampal neurons that selectively fire or cease to fire when the mouse perceives nests or beds, regardless of their locations and environments. Parametric analyses show that responses of nest cells remain invariant over changes in the nests' physical shape, style, color, odor, or construction materials; rather, their responses are driven by conscious awareness and physical determination of the categorical features that would functionally define nests. Such functionality-based abstraction and generalization of conceptual knowledge, emerging from episodic experiences, suggests that the hippocampus is an intrinsic part of the hierarchical structure for generating concepts and knowledge in the brain.Here is one figure and one movie from the paper:
Legend - Invariant responses over the geometric shapes, physical appearances, colors, construction materials, etc. (A) Invariant responses of Cell #1 to the geometric shapes of nests. As shown by both the perievent spike rasters and perievent spike histograms, Cell #1 exhibited a significant firing increase in response to a new circular nest made out of the top part of a cardboard coffee cup with a wall height of 2.5 cm and a diameter of 7.5 cm (Left), a square cardboard nest (Center), and a triangular cardboard nest (Right). (B) Invariant responses of Cell #1 to nests made from different materials. A metal nest (Left), plastic nest (Center), and porcelain nest (Right) were tested. (C) The cell also increased its firing when the mouse encountered natural cotton nests (Left and Center) but not to five cotton balls that were simply lumped together (Right). The bin width in the perievent spike histogram is 250 ms.
Click to download Movie 1. Description of movie: "Transient-on" type of nest cell responses and its functionality-based encoding of conceptual knowledge of nests. The first movie segment shows that Cell #1 of mouse-A exhibited "transient-on"-type responses to the home nest but not to another similarly shaped, smaller circular object (water cup). The second movie segment shows that Cell #1 did not respond to the plastic nest that was placed in an inverted manner (so that it would function as a small stage). However, once the plastic nest reverted back to its normal nest position, the cell exhibited robust firing up to 40 Hz. Because it is the same object and placed at the same location, this inversion experiment demonstrates that the nest cell encodes the functionality of the nest rather than merely physical appearances, materials, or spatial location, etc.

Why the rich behave badly... "disinhibition"
 Richard Conniff writes an OpEd piece in the 4/4/07 NY Times (PDF here) pointing out the work of UC Berkely psychologist Dacher Keltner and others who study the effect of power on social perception and behavior. Their work suggests that elevated power leads to behavioral disinhibition and reduced vigilance. They find that ideological partisans with power construe their dispute in more stereotypical, polarized fashion, that elevated social status leads to disinhibited social behavior, and that power, whether derived from group status or experimental manipulation, relates to the experience of increased positive emotion and reduced negative emotion.
Richard Conniff writes an OpEd piece in the 4/4/07 NY Times (PDF here) pointing out the work of UC Berkely psychologist Dacher Keltner and others who study the effect of power on social perception and behavior. Their work suggests that elevated power leads to behavioral disinhibition and reduced vigilance. They find that ideological partisans with power construe their dispute in more stereotypical, polarized fashion, that elevated social status leads to disinhibited social behavior, and that power, whether derived from group status or experimental manipulation, relates to the experience of increased positive emotion and reduced negative emotion.In what Conniff calls the "Cookie Monster Experiment" (a much milder version of the Stanford Prison Experiment) Keltner:
...took groups of three ordinary volunteers and randomly put one of them in charge. Each trio had a half-hour to work through a boring social survey. Then a researcher came in and left a plateful of precisely five cookies. Care to guess which volunteer typically grabbed an extra cookie? The volunteer who had randomly been assigned the power role was also more likely to eat it with his mouth open, spew crumbs on partners and get cookie detritus on his face and on the table.Keltner theorizes:
It reminded the researchers of powerful people they had known in real life. One of them, for instance, had attended meetings with a magazine mogul who ate raw onions and slugged vodka from the bottle, but failed to share these amuse-bouches with his guests. Another had been through an oral exam for his doctorate at which one faculty member not only picked his ear wax, but held it up to dandle lovingly in the light.
...that getting power causes people to focus so keenly on the potential rewards, like money, sex, public acclaim or an extra chocolate-chip cookie — not necessarily in that order, or frankly, any order at all, but preferably all at once — that they become oblivious to the people around them.
Indeed, the people around them may abet this process, since they are often subordinates intent on keeping the boss happy. So for the boss, it starts to look like a world in which the traffic lights are always green (and damn the pedestrians). Professor Keltner and his fellow researchers describe it as an instance of “approach/inhibition theory” in action: As power increases, it fires up the behavioral approach system and shuts down behavioral inhibition.
Tuesday, April 10, 2007
Rapid memory consolidation shown...
Memory encoding occurs rapidly, and requires the hippocampus, but the consolidation of memory in the neocortex has long been held to be a more gradual process. Tse et al now show in experiments with rats learning the position of new treats placed in a maze:

...that systems consolidation can occur extremely quickly if an associative "schema" into which new information is incorporated has previously been created. In experiments using a hippocampal-dependent paired-associate task for rats, the memory of flavor-place associations became persistent over time as a putative neocortical schema gradually developed. New traces, trained for only one trial, then became assimilated and rapidly hippocampal-independent. Schemas also played a causal role in the creation of lasting associative memory representations during one-trial learning. The concept of neocortical schemas may unite psychological accounts of knowledge structures with neurobiological theories of systems memory consolidation. (PDF here)And, from Larry Squire's perspective article on this paper (PDF here):

Legend. (click on figure to enlarge) When a rat learns associations between flavors and spatial locations, as studied by Tse et al. (1), the associations are initially learned as individual facts (left). With extended training, the animal develops an organized structure or schema for flavors and places (middle). This organized knowledge structure (bold lines) can then support rapid learning of new associations in a single trial and the rapid consolidation of information into the neocortex (right).
Do animals experience past and future?
Carl Zimmer writes a nice piece in the 4/3 NYTimes on "Time in the Animal Mind" (PDF HERE).
 Legend: Scrub jays, left, seem able to plan for the future in experiments, hiding today’s pine nuts for tomorrow’s breakfast. Squirrel monkeys also seem to think about future consequences, while hummingbirds seem to recall time and location of visits to flowers, and rats to remember where they encountered food in a maze. (Credits: from left, Adam Jones/Photo Researchers; Luke MacGregor/Reuters; Esteban Felix/Associated Press; Will & Deni McIntyre/Photo Researchers)
Legend: Scrub jays, left, seem able to plan for the future in experiments, hiding today’s pine nuts for tomorrow’s breakfast. Squirrel monkeys also seem to think about future consequences, while hummingbirds seem to recall time and location of visits to flowers, and rats to remember where they encountered food in a maze. (Credits: from left, Adam Jones/Photo Researchers; Luke MacGregor/Reuters; Esteban Felix/Associated Press; Will & Deni McIntyre/Photo Researchers)
The animals shown in the figure have been shown to have impressive powers of memory, but this doesn't have to imply having a sense of memory or self, i.e. 'thinking about' past or future.
Some argue that mental time travel is distinctive to hominids. Humans can remember events long past and envision the future, and recent experiments by Schacter's lab at Harvard have shown that brain areas involved in episodic memory become active also when people think of themselves in the future.
What about animal's sense of the future? Can they plan ahead? Nicola, Univ. of Cambridge, has done interesting experiments showing sophisticated memory in scrub jays, and she
 Legend: Scrub jays, left, seem able to plan for the future in experiments, hiding today’s pine nuts for tomorrow’s breakfast. Squirrel monkeys also seem to think about future consequences, while hummingbirds seem to recall time and location of visits to flowers, and rats to remember where they encountered food in a maze. (Credits: from left, Adam Jones/Photo Researchers; Luke MacGregor/Reuters; Esteban Felix/Associated Press; Will & Deni McIntyre/Photo Researchers)
Legend: Scrub jays, left, seem able to plan for the future in experiments, hiding today’s pine nuts for tomorrow’s breakfast. Squirrel monkeys also seem to think about future consequences, while hummingbirds seem to recall time and location of visits to flowers, and rats to remember where they encountered food in a maze. (Credits: from left, Adam Jones/Photo Researchers; Luke MacGregor/Reuters; Esteban Felix/Associated Press; Will & Deni McIntyre/Photo Researchers)The animals shown in the figure have been shown to have impressive powers of memory, but this doesn't have to imply having a sense of memory or self, i.e. 'thinking about' past or future.
Some argue that mental time travel is distinctive to hominids. Humans can remember events long past and envision the future, and recent experiments by Schacter's lab at Harvard have shown that brain areas involved in episodic memory become active also when people think of themselves in the future.
What about animal's sense of the future? Can they plan ahead? Nicola, Univ. of Cambridge, has done interesting experiments showing sophisticated memory in scrub jays, and she
"recently tested her scrub jays for foresight. She and her colleagues put the birds in three adjoining compartments for six days. Each morning the birds were shut for two hours in one of two rooms. In one room they got nothing to eat. In the other room, they got powdered pine nuts (the scrub jays can eat the powder, but they cannot cache it). For the rest of the day, each bird could move around all three rooms and enjoy more powdered nuts.This and similar experiments in other animals may over the next few years provide more compelling evidence that animals do plan ahead, and thus take away yet another feature that many have thought distinctive to humans.
On the seventh day, the scientists switched the powdered pine nuts with real ones. If the birds were so inclined, they could cache the pine nuts in ice cube trays the scientists put in the two morning rooms. “If I’m a bird, what I could do is take some of the provisions and hide it in there so that if I do wake up there in the morning, I can get my own breakfast,” Dr. Clayton said.
Dr. Clayton found that the birds put over three times more pine nuts in the no-breakfast room than in the breakfast room. She argues that the results mean that birds can take action for their future needs, knowing what they’ll need and where they’ll need it."
Monday, April 09, 2007
Friday, April 06, 2007
MindBlog leaves 'paradise'
Postings may become erratic or non-existent for several days as I leave my snowbird site, Fort Lauderdale, and drive back to Madison Wisconsin on Monday, April 9.
This is what I leave behind: The beach and the condo:


This is what I leave behind: The beach and the condo:


Motor cortex for the hand and numerical counting
The April 2007 issue of the Journal of Cognitive Neuroscience has interesting articles on this topic by Andres et al. and Sato et al. Their abstracts:
Andres et al.
Sato et al.
Andres et al.
The finding that number processing activates a cortical network partly overlapping that recruited for hand movements has renewed interest in the relationship between number and finger representations. Further evidence about a possible link between fingers and numbers comes from developmental studies showing that finger movements play a crucial role in learning counting. However, increased activity in hand motor circuits during counting may unveil unspecific processes, such as shifting attention, reciting number names, or matching items with a number name. To address this issue, we used transcranial magnetic stimulation to measure changes in corticospinal (CS) excitability during a counting task performed silently and using either numbers or letters of the alphabet to enumerate items. We found an increased CS excitability of hand muscles during the counting task, irrespective of the use of numbers or letters, whereas it was unchanged in arm and foot muscles. Control tasks allowed us to rule out a possible influence of attention allocation or covert speech on CS excitability increase of hand muscles during counting. The present results support a specific involvement of hand motor circuits in counting because no CS changes were found in arm and foot muscles during the same task. However, the contribution of hand motor areas is not exclusively related to number processing because an increase in CS excitability was also found when letters were used to enumerate items. This finding suggests that hand motor circuits are involved whenever items have to be put in correspondence with the elements of any ordered series.
Sato et al.
Developmental and cross-cultural studies show that finger counting represents one of the basic number learning strategies. However, despite the ubiquity of such an embodied strategy, the issue of whether there is a neural link between numbers and fingers in adult, literate individuals remains debated. Here, we used transcranial magnetic stimulation to study changes of excitability of hand muscles of individuals performing a visual parity judgment task, a task not requiring counting, on Arabic numerals from 1 to 9. Although no modulation was observed for the left hand muscles, an increase in amplitude of motor-evoked potentials was found for the right hand muscles. This increase was specific for smaller numbers (1 to 4) as compared to larger numbers (6 to 9). These findings indicate a close relationship between hand/finger and numerical representations.
Thursday, April 05, 2007
Most popular consciousness papers for March
Again I pass on the list of articles downloaded most frequently from the ASSC Eprints Archive
The "five most popular papers" are:
1. Windt, Jennifer Michelle and Metzinger, Thomas (2006) *The philosophy of
dreaming and self-consciousness: What happens to the experiential subject
during the dream state?* In: The new science of dreaming. Praeger
Imprint/Greenwood Publishers, Estport, CT. With 1488 downloads from 23
countries. See:http://eprints.assc.caltech.edu/200/
2. Sagiv, Noam and Ward, Jamie (2006) *Crossmodal interactions: lessons from
synesthesia.* In: Visual Perception, Part 2 - Fundamentals of Awareness:
Multi-Sensory Integration and High-Order Perception. Progress in Brain
Research, Volume 155. Elsevier, pp. 259-271. ISBN 0444519270. With 1034
downloads from 17 countries. See: http://eprints.assc.caltech.edu/224/
3. Carruthers, Peter (2007) *The illusion of conscious will.* In: Synthese,
96. With 879 downloads from 19 countries. See: http://eprints . assc
.caltech.edu/213/
4. Robbins, Stephen E (2006) *Bergson and the holographic theory of
mind.* Phenomenology
and the Cognitive Sciences, 5. pp. 365-394. With 760 downloads from 16
countries. See:http://eprints.assc.caltech.edu/206/
5. Chai-Youn, Kim and Blake, Randolph (2005) *Psychophysical magic:
rendering the visible 'invisible'.* Trends in Cognitive Science, 9 (8).
pp. 381-8. With 729 downloads from 15 countries. See:
http://eprints.assc.caltech.edu/30/.
The "five most popular papers" are:
1. Windt, Jennifer Michelle and Metzinger, Thomas (2006) *The philosophy of
dreaming and self-consciousness: What happens to the experiential subject
during the dream state?* In: The new science of dreaming. Praeger
Imprint/Greenwood Publishers, Estport, CT. With 1488 downloads from 23
countries. See:http://eprints.assc.caltech.edu/200/
2. Sagiv, Noam and Ward, Jamie (2006) *Crossmodal interactions: lessons from
synesthesia.* In: Visual Perception, Part 2 - Fundamentals of Awareness:
Multi-Sensory Integration and High-Order Perception. Progress in Brain
Research, Volume 155. Elsevier, pp. 259-271. ISBN 0444519270. With 1034
downloads from 17 countries. See: http://eprints.assc.caltech.edu/224/
3. Carruthers, Peter (2007) *The illusion of conscious will.* In: Synthese,
96. With 879 downloads from 19 countries. See: http://eprints . assc
.caltech.edu/213/
4. Robbins, Stephen E (2006) *Bergson and the holographic theory of
mind.* Phenomenology
and the Cognitive Sciences, 5. pp. 365-394. With 760 downloads from 16
countries. See:http://eprints.assc.caltech.edu/206/
5. Chai-Youn, Kim and Blake, Randolph (2005) *Psychophysical magic:
rendering the visible 'invisible'.* Trends in Cognitive Science, 9 (8).
pp. 381-8. With 729 downloads from 15 countries. See:
http://eprints.assc.caltech.edu/30/.
Self and social cognition in the brain
Uddin et al. speculate in Trends in Cognitive Science (Volume 11, Issue 4 , April 2007, Pages 153-157) about the respective roles of cortical midline structures (CMS) and frontoparietal mirror neurons (MNS) in evaluation of self and others. Below I relay one of their summary paragraphs, and the PDF which contains images of relevant brain regions can be obtained HERE.
It has recently been proposed that internally oriented processes that focus on one's own or others’ mental states rely on cortical midline structures, whereas externally focused processes based on one's own or others’ visible features and actions rely on lateral frontoparietal networks. We suggest here a similar distinction, which might further reconcile disparate findings with regard to the various proposed functions of cortical midline structures, while incorporating what is known about the role of the human mirror-neuron system in social cognition. Whereas there is mounting evidence that the right frontoparietal system is involved in representing the physical, embodied self (in addition to its role in understanding the actions of others), the cortical midline structures that comprise the default-mode network seem to be more involved in maintaining a self-representation in evaluative terms, which requires self-referential processing and understanding of others’ mental states. We speculate that the CMS might support evaluative simulation in the same way that the MNS supports motor simulation. This distinction serves as a practical division of labor between two networks that are specialized for two related processes that are crucial to navigating the social world. The mirror-neuron system provides the essential physical other-to-self mapping that is necessary for comprehending physical actions of intentional agents, whereas cortical midline structures maintain and support processes that are related to understanding complex psychological aspects of others, such as attitudes, perhaps by simulation of one's own attitudes.
Wednesday, April 04, 2007
Emotion and consciousness - a Review
Tsuchiya and Adolphs at Cal. Tech. offer a broad review of this topic in Trends in Cognitive Sciences (Volume 11, Issue 4, April 2007, Pages 158-167) which has some excellent illustrations for teaching. Here I show the abstract and three figures from the article, the PDF is HERE.
 Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html).
Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html).
 Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].
Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].
 Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].
Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].
 Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html).
Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html). Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].
Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].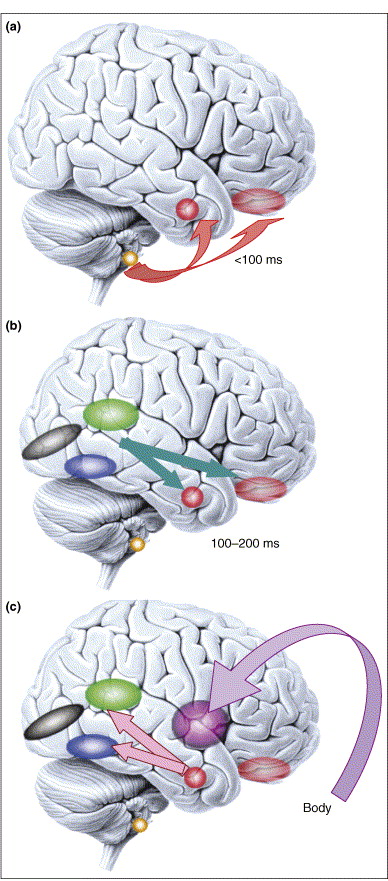 Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].
Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].
Rhythms linking episodic and procedural memory systems.
I pass on this abstract from an interesting paper by DeCoteau et al. PDF is HERE.
The striatum and hippocampus are conventionally viewed as complementary learning and memory systems, with the hippocampus specialized for fact-based episodic memory and the striatum for procedural learning and memory. Here we directly tested whether these two systems exhibit independent or coordinated activity patterns during procedural learning. We trained rats on a conditional T-maze task requiring navigational and cue-based associative learning. We recorded local field potential (LFP) activity with tetrodes chronically implanted in the caudoputamen and the CA1 field of the dorsal hippocampus during 6–25 days of training. We show that simultaneously recorded striatal and hippocampal theta rhythms are modulated differently as the rats learned to perform the T-maze task but nevertheless become highly coherent during the choice period of the maze runs in rats that successfully learned the task. Moreover, in the rats that acquired the task, the phase of the striatal–hippocampal theta coherence was modified toward a consistent antiphase relationship, and these changes occurred in proportion to the levels of learning achieved. We suggest that rhythmic oscillations, including theta-band activity, could influence not only neural processing in cortico-basal ganglia circuits but also dynamic interactions between basal ganglia-based and hippocampus-based forebrain circuits during the acquisition and performance of learned behaviors. Experience-dependent changes in coordination of oscillatory activity across brain structures thus may parallel the well known plasticity of spike activity that occurs as a function of experience.
Tuesday, April 03, 2007
Exercise increases nerve cell growth in brain area crucial to cognitive aging.
Pereira et al. have used an interesting approach to demonstrate that exercise causes the growth of new nerve cells in the dentate gyrus (part of the hippocampus) of mice and humans. Because new nerve cell formation usually is accompanied by synthesis of new blood vessels and increased blood flow, they used MRI measurement of cerebral blood volume as an assay of new nerve cell formation in the living brain. Here is their abstract, and a figure from the paper. The PDF of the paper can be downloaded HERE.
With continued debate over the functional significance of adult neurogenesis, identifying an in vivo correlate of neurogenesis has become an important goal. Here we rely on the coupling between neurogenesis and angiogenesis and test whether MRI measurements of cerebral blood volume (CBV) provide an imaging correlate of neurogenesis. First, we used an MRI approach to generate CBV maps over time in the hippocampal formation of exercising mice. Among all hippocampal subregions, exercise was found to have a primary effect on dentate gyrus CBV, the only subregion that supports adult neurogenesis. Moreover, exercise-induced increases in dentate gyrus CBV were found to correlate with postmortem measurements of neurogenesis. Second, using similar MRI technologies, we generated CBV maps over time in the hippocampal formation of exercising humans. As in mice, exercise was found to have a primary effect on dentate gyrus CBV, and the CBV changes were found to selectively correlate with cardiopulmonary and cognitive function. Taken together, these findings show that dentate gyrus CBV provides an imaging correlate of exercise-induced neurogenesis and that exercise differentially targets the dentate gyrus, a hippocampal subregion important for memory and implicated in cognitive aging.Legend - (click on figure to enlarge) Exercise selectively increases dentate gyrus CBV in humans. (a) Exercise had a selective effect on dentate gyrus CBV. Bar graph shows the mean relative CBV (rCBV) values for each hippocampal subregion before exercise (open bars) and after exercise (filled bars). As in mice, the dentate gyrus was the only hippocampal subregion that showed a significant exercise effect, whereas the entorhinal cortex showed a nonsignificant increase in CBV. (b) An individual example. (Left) High-resolution MRI slice that visualizes the external morphology and internal architecture of the hippocampal formation. (Center) Parcellation of the hippocampal subregions (green, entorhinal cortex; red, dentate gyrus; blue, CA1 subfield; yellow, subiculum). (Right) Hippocampal CBV map (warmer colors reflect higher CBV).

Mirror Neurons - gee whiz popular video
A engaging and easy to watch popularized version of material I've written about earlier (see 1/20/07; 12/19/06; and my lecture 10/27/06)).
Monday, April 02, 2007
Social exclusion causes an empathy block.
From the Editor's choice section of the March 23 issue of Science Magazine, a discussion of Twenge et al., in the Journal of Personality & Social Psychology. 92(1):56-66, January 2007:
Everyday experience confirms the general belief that humans are social animals; the neural pathways subserving prosocial behaviors are a subject of current research, and the evolutionary origins of these behaviors are hotly debated. Although there is evidence that social exclusion can elicit redoubled efforts to develop social connections, the consequences of exclusion are predominantly negative--feeling hurt, acting belligerently, or adopting a lone-wolf lifestyle--and Twenge et al. have begun to examine what might mediate these apparently atypical responses.Here is the Twenge et al. abstract, the PDF can be downloaded HERE.
Using a variety of experimental contexts (such as the canonical spilled-pencils incident) and measures (such as donations of money or cooperation in a prisoner's dilemma game), they find that being characterized as having a high likelihood of a prosocial lifestyle with many strong relationships, such as marriage, resulted in participants helping to pick up pencils (on average, 8 out of 20 spilled) versus the performance of those labeled as being apt to lead solitary lives (less than 1 pencil picked up). As to what factors mediate the extent (or absence) of prosocial behavior, some of the likely candidates (trusting the other or having a sense of belonging) did not register, whereas empathic concern did. Combining this finding with an earlier one, which showed that social exclusion activates the neural circuits encoding pain, produces the speculation that an after-effect of rejection is an emotional numbness or an inability to mirror the affective states of others.
In 7 experiments, the authors manipulated social exclusion by telling people that they would end up alone later in life or that other participants had rejected them. Social exclusion caused a substantial reduction in prosocial behavior. Socially excluded people donated less money to a student fund, were unwilling to volunteer for further lab experiments, were less helpful after a mishap, and cooperated less in a mixed-motive game with another student. The results did not vary by cost to the self or by recipient of the help, and results remained significant when the experimenter was unaware of condition. The effect was mediated by feelings of empathy for another person but was not mediated by mood, state self-esteem, belongingness, trust, control, or self-awareness. The implication is that rejection temporarily interferes with emotional responses, thereby impairing the capacity for empathic understanding of others, and as a result, any inclination to help or cooperate with them is undermined.
Neurite Outgrowth
I've always been fascinated by time lapse videos of neurons growing on the surface of a culture dish. Here is one example:
Subscribe to:
Posts (Atom)