It has recently been proposed that internally oriented processes that focus on one's own or others’ mental states rely on cortical midline structures, whereas externally focused processes based on one's own or others’ visible features and actions rely on lateral frontoparietal networks. We suggest here a similar distinction, which might further reconcile disparate findings with regard to the various proposed functions of cortical midline structures, while incorporating what is known about the role of the human mirror-neuron system in social cognition. Whereas there is mounting evidence that the right frontoparietal system is involved in representing the physical, embodied self (in addition to its role in understanding the actions of others), the cortical midline structures that comprise the default-mode network seem to be more involved in maintaining a self-representation in evaluative terms, which requires self-referential processing and understanding of others’ mental states. We speculate that the CMS might support evaluative simulation in the same way that the MNS supports motor simulation. This distinction serves as a practical division of labor between two networks that are specialized for two related processes that are crucial to navigating the social world. The mirror-neuron system provides the essential physical other-to-self mapping that is necessary for comprehending physical actions of intentional agents, whereas cortical midline structures maintain and support processes that are related to understanding complex psychological aspects of others, such as attitudes, perhaps by simulation of one's own attitudes.
Thursday, April 05, 2007
Self and social cognition in the brain
Uddin et al. speculate in Trends in Cognitive Science (Volume 11, Issue 4 , April 2007, Pages 153-157) about the respective roles of cortical midline structures (CMS) and frontoparietal mirror neurons (MNS) in evaluation of self and others. Below I relay one of their summary paragraphs, and the PDF which contains images of relevant brain regions can be obtained HERE.
Wednesday, April 04, 2007
Emotion and consciousness - a Review
Tsuchiya and Adolphs at Cal. Tech. offer a broad review of this topic in Trends in Cognitive Sciences (Volume 11, Issue 4, April 2007, Pages 158-167) which has some excellent illustrations for teaching. Here I show the abstract and three figures from the article, the PDF is HERE.
 Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html).
Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html).
 Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].
Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].
 Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].
Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].
 Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html).
Figure 2. (click to enlarge) Brain regions that are important for emotion state (blue), feeling of emotion (red) and level of consciousness (green). Other components of an emotion state, and the content of consciousness, are presumed to rely on more variable and distributed structures that would depend on the particular kind of emotion or conscious experience and, therefore, are not depicted here. (a) Sagittal view shows that the hypothalamus, amygdala, brainstem nuclei, including periaquedactal gray and parabrachial nuclei, orbitofrontal cortex and anterior cingulate cortex are important for the expression of emotion (blue). Anterior and posterior cingulate, including precuneus, and orbitofrontal cortex are important for the experience of emotion (red). Intralaminar thalamus and the ascending reticular formation are necessary for the maintenance of arousal and wakefulness – that is, the level (state) of consciousness 8 and 12. The intralaminar thalamic nucleus is a diffuse structure, enlarged here for illustration purposes. (b) Coronal slice at the level of the white line in (a). The thalamus, hypothalamus and amygdala are shown again. Insular cortex (red) is an important structure for the experience of emotion [also shown in (d)]. (c) Bilateral prefrontal and parietal cortices are broadly important for the level of consciousness (green) [36]. The figure omits other important central nervous system components of emotion, such as the rostral ventrolateral medulla (important for control of autonomic function) and components of the spinal cord itself, all of which contribute to substantial processing that is related to interoceptive and homeostatic information [46], and also parts of the nucleus accumbens and ventral pallidum that participate in reward and positive affect 4 and 60 (See also Figure 3c). (d) The surface of the prefrontal cortex has been removed to reveal the insular cortex (red). Human brain images are adapted, with permission, from the Digital Anatomist Project at the University of Washington (http://www9.biostr.washington.edu/da.html). Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].
Figure 3. (click to enlarge) Shared neural substrates for emotion and consciousness. (a) Data from functional imaging show that a frontoparietal network is compromised in coma, vegetative states, sleep and under anesthesia (black regions; but also see green regions in Figure 2). Abbreviations: F, prefrontal; MF, mesiofrontal; P, posterior parietal; Pr, posterior cingulate/precuneus. (a) reproduced, with permission, from Ref. [8]. (b) In a quiet but awake resting state, there are two distinct networks of structures in the human brain that are either correlated (color-coded red to orange) or anti-correlated (blue to green) with the parietal cortex, indicated by the arrow. The blue–green network of structures might be particularly important for reflective and interoceptive processing and crucial to the level of consciousness and feeling emotions. (b) reproduced, with permission, from Ref. [14]. (c) Anatomical structures that are important for feelings (emotion experience) involve homeostatic loops. A main circuit for efferent autonomic control arises from distributed cortical sectors in anterior and subgenual cingulate cortex, medial orbital cortex and insula, as well as from amygdala. These structures in turn project to paraventricular hypothalamus and periaqueductal gray (PAG) matter, from there to integrative centers in the medulla (RVLM, rostral ventrolateral medulla; VMM, ventromedial medulla) and the spinal cord, and then out to effector organs. Afferent autonomic processing occurs in part through dedicated sensory channels and involves brainstem nuclei, including the parabrachial nucleus (PB), the hypothalamus, thalamus (VM, ventromedial nucleus), and then again the same set of cortical regions. With the exception of the thalamus, there are homeostatic control loops at multiple levels, involving all structures in both afference and efference. Many of these structures are activated in a variety of experiments that involve emotion experiences 16, 17 and 46, making them promising candidates for the neural correlates of emotion experience 3 and 46. It has been proposed that primates possess a unique mapping of autonomic interoceptive information within the insular cortex that forms the substrate of conscious feelings [46]. The afferent limb is shown in the top row and the efferent limb in the bottom row; please note that only a subset of the connectivity is depicted in this figure for clarity. The red lines indicate pathways in primates thought to be more phylogenetically recent that provide a direct thalamocortical input, reflecting the physiological condition of the body. Abbreviations: A1, catecholaminergic cell groups A1; ACC, anterior cingulate; MD, medial dorsal nucleus; NTS, nucleus of the solitary tract. (c) modified, with permission, from Ref. [72].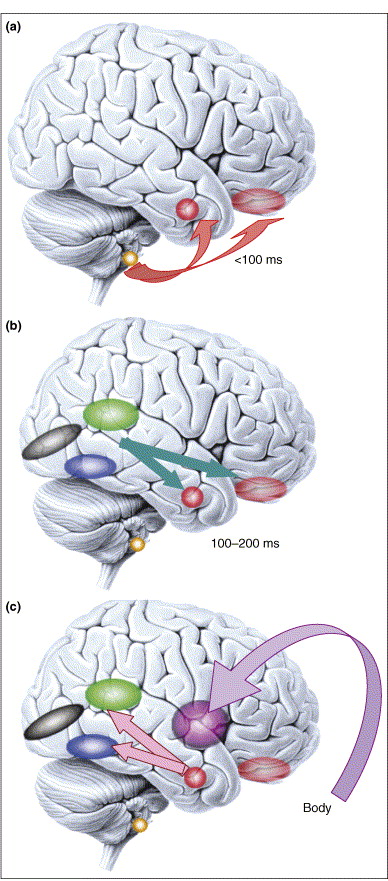 Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].
Figure 4. (click to enlarge) Microgenesis of emotional processing. Emotional responses span a large temporal range (from 100 ms or less, to minutes). (a) Responses to emotional visual stimuli can occur rapidly in prefrontal cortex [50] or amygdala, in part mediated by subcortical inputs. Emotional response in the amygdala also influences early visual processing [51] and is modulated by volitional self-regulation 47 and 52. (b) At later time slices (100–200 ms), sensory cortices provide more detailed input to emotion-inducing structures like the amygdala. Two components that are important to face processing are shown: the superior temporal cortex (green), important for encoding dynamic information such as facial expression, and the fusiform gyrus (blue), important for encoding static information such as identity. (c) Once the emotional meaning of a stimulus has been evaluated by the brain, emotional responses are triggered in the body via projections from amygdala and medial prefrontal cortex to brainstem nuclei and hypothalamus (not shown), and are in turn represented in structures such as the insula. This figure emphasizes that what we refer to as an ‘emotion state’ throughout this article is in fact a complex set of processes that unfold at various points in time. Color key: black, primary visual cortex; blue, fusiform gyrus; green, superior temporal cortex; purple, insula; faint red, orbitofrontal cortex; solid red, amygdala; yellow, superior colliculus. Reproduced, with permission, from Ref. [73].
Rhythms linking episodic and procedural memory systems.
I pass on this abstract from an interesting paper by DeCoteau et al. PDF is HERE.
The striatum and hippocampus are conventionally viewed as complementary learning and memory systems, with the hippocampus specialized for fact-based episodic memory and the striatum for procedural learning and memory. Here we directly tested whether these two systems exhibit independent or coordinated activity patterns during procedural learning. We trained rats on a conditional T-maze task requiring navigational and cue-based associative learning. We recorded local field potential (LFP) activity with tetrodes chronically implanted in the caudoputamen and the CA1 field of the dorsal hippocampus during 6–25 days of training. We show that simultaneously recorded striatal and hippocampal theta rhythms are modulated differently as the rats learned to perform the T-maze task but nevertheless become highly coherent during the choice period of the maze runs in rats that successfully learned the task. Moreover, in the rats that acquired the task, the phase of the striatal–hippocampal theta coherence was modified toward a consistent antiphase relationship, and these changes occurred in proportion to the levels of learning achieved. We suggest that rhythmic oscillations, including theta-band activity, could influence not only neural processing in cortico-basal ganglia circuits but also dynamic interactions between basal ganglia-based and hippocampus-based forebrain circuits during the acquisition and performance of learned behaviors. Experience-dependent changes in coordination of oscillatory activity across brain structures thus may parallel the well known plasticity of spike activity that occurs as a function of experience.
Tuesday, April 03, 2007
Exercise increases nerve cell growth in brain area crucial to cognitive aging.
Pereira et al. have used an interesting approach to demonstrate that exercise causes the growth of new nerve cells in the dentate gyrus (part of the hippocampus) of mice and humans. Because new nerve cell formation usually is accompanied by synthesis of new blood vessels and increased blood flow, they used MRI measurement of cerebral blood volume as an assay of new nerve cell formation in the living brain. Here is their abstract, and a figure from the paper. The PDF of the paper can be downloaded HERE.
With continued debate over the functional significance of adult neurogenesis, identifying an in vivo correlate of neurogenesis has become an important goal. Here we rely on the coupling between neurogenesis and angiogenesis and test whether MRI measurements of cerebral blood volume (CBV) provide an imaging correlate of neurogenesis. First, we used an MRI approach to generate CBV maps over time in the hippocampal formation of exercising mice. Among all hippocampal subregions, exercise was found to have a primary effect on dentate gyrus CBV, the only subregion that supports adult neurogenesis. Moreover, exercise-induced increases in dentate gyrus CBV were found to correlate with postmortem measurements of neurogenesis. Second, using similar MRI technologies, we generated CBV maps over time in the hippocampal formation of exercising humans. As in mice, exercise was found to have a primary effect on dentate gyrus CBV, and the CBV changes were found to selectively correlate with cardiopulmonary and cognitive function. Taken together, these findings show that dentate gyrus CBV provides an imaging correlate of exercise-induced neurogenesis and that exercise differentially targets the dentate gyrus, a hippocampal subregion important for memory and implicated in cognitive aging.Legend - (click on figure to enlarge) Exercise selectively increases dentate gyrus CBV in humans. (a) Exercise had a selective effect on dentate gyrus CBV. Bar graph shows the mean relative CBV (rCBV) values for each hippocampal subregion before exercise (open bars) and after exercise (filled bars). As in mice, the dentate gyrus was the only hippocampal subregion that showed a significant exercise effect, whereas the entorhinal cortex showed a nonsignificant increase in CBV. (b) An individual example. (Left) High-resolution MRI slice that visualizes the external morphology and internal architecture of the hippocampal formation. (Center) Parcellation of the hippocampal subregions (green, entorhinal cortex; red, dentate gyrus; blue, CA1 subfield; yellow, subiculum). (Right) Hippocampal CBV map (warmer colors reflect higher CBV).

Mirror Neurons - gee whiz popular video
A engaging and easy to watch popularized version of material I've written about earlier (see 1/20/07; 12/19/06; and my lecture 10/27/06)).
Monday, April 02, 2007
Social exclusion causes an empathy block.
From the Editor's choice section of the March 23 issue of Science Magazine, a discussion of Twenge et al., in the Journal of Personality & Social Psychology. 92(1):56-66, January 2007:
Everyday experience confirms the general belief that humans are social animals; the neural pathways subserving prosocial behaviors are a subject of current research, and the evolutionary origins of these behaviors are hotly debated. Although there is evidence that social exclusion can elicit redoubled efforts to develop social connections, the consequences of exclusion are predominantly negative--feeling hurt, acting belligerently, or adopting a lone-wolf lifestyle--and Twenge et al. have begun to examine what might mediate these apparently atypical responses.Here is the Twenge et al. abstract, the PDF can be downloaded HERE.
Using a variety of experimental contexts (such as the canonical spilled-pencils incident) and measures (such as donations of money or cooperation in a prisoner's dilemma game), they find that being characterized as having a high likelihood of a prosocial lifestyle with many strong relationships, such as marriage, resulted in participants helping to pick up pencils (on average, 8 out of 20 spilled) versus the performance of those labeled as being apt to lead solitary lives (less than 1 pencil picked up). As to what factors mediate the extent (or absence) of prosocial behavior, some of the likely candidates (trusting the other or having a sense of belonging) did not register, whereas empathic concern did. Combining this finding with an earlier one, which showed that social exclusion activates the neural circuits encoding pain, produces the speculation that an after-effect of rejection is an emotional numbness or an inability to mirror the affective states of others.
In 7 experiments, the authors manipulated social exclusion by telling people that they would end up alone later in life or that other participants had rejected them. Social exclusion caused a substantial reduction in prosocial behavior. Socially excluded people donated less money to a student fund, were unwilling to volunteer for further lab experiments, were less helpful after a mishap, and cooperated less in a mixed-motive game with another student. The results did not vary by cost to the self or by recipient of the help, and results remained significant when the experimenter was unaware of condition. The effect was mediated by feelings of empathy for another person but was not mediated by mood, state self-esteem, belongingness, trust, control, or self-awareness. The implication is that rejection temporarily interferes with emotional responses, thereby impairing the capacity for empathic understanding of others, and as a result, any inclination to help or cooperate with them is undermined.
Neurite Outgrowth
I've always been fascinated by time lapse videos of neurons growing on the surface of a culture dish. Here is one example:
The Male-Warrior Hypothesis
Some clips from Van Vugt et al:
Evolutionary scientists argue that human cooperation is the product of a long history of competition among rival groups. There are various reasons to believe that this logic applies particularly to men. In three experiments, using a step-level public-goods task, we found that men contributed more to their group if their group was competing with other groups than if there was no intergroup competition. Female cooperation was relatively unaffected by intergroup competition. These findings suggest that men respond more strongly than women to intergroup threats. ..These findings fit nicely with an evolutionary hypothesis about specific male intergroup adaptations—the male-warrior hypothesis—and such evolved intergroup traits are likely to be reinforced through cultural processes, for example, during childhood socialization...Women's social psychology is likely to be shaped more strongly by different kinds of needs, such as defending their offspring and creating supportive social networksThe article's PDF can be downloaded HERE.
Friday, March 30, 2007
Looking for hidden signs of consciousness
In the 22 March issue of Nature, Kerri Smith discusses the debate over a 'brain-activity' test for patients in a vegetative state (PDF download HERE).
 Imagining spatial navigation (left) and playing tennis.
Imagining spatial navigation (left) and playing tennis.
A team, led by Adrian Owen of the MRC Cognition and Brain Sciences Unit in Cambridge, UK, used functional magnetic resonance imaging (fMRI) to show that a woman left in a vegetative state after a car accident could respond to requests to imagine playing tennis or navigate around her house (A. Owen et al. Science 313, 1402; 2006)... Laureys, a member of this team, has now tested this technique on 24 healthy volunteers, who were similarly instructed to imagine either walking around their house or playing tennis. The tasks activate separate networks in the brain, and the scans proved able to tell correctly which task was being performed (M. Boly et al. NeuroImage doi:10.1016/j.neuroimage.2007.02.047; 2007)...showing that the method works reliably in healthy brains proves its robustness. "Our challenge is to find markers that tell us 'this is a hopeless case' or 'this is a case where we should increase our therapeutic efforts'," says Laureys.
 Imagining spatial navigation (left) and playing tennis.
Imagining spatial navigation (left) and playing tennis.
A primer on, and reservations about, TMS - transcranial magnetic stimulation
O'Shea and Walsh offer a brief introduction to TMS, and its use in cognitive neuroscience:
There is controversy over the ethics of using this techniques on humans, it can cause seizures or psychosis in some subjects. This is the subject of a letter to the editor in the March 23 issue of Science from Leslie Sargent Jones of the Univ. of S. Carolina:
As any schoolboy with a toolkit or a broken toy soon appreciates, to find out how a machine works you need to take it apart, and to put it back together again, you need to know how it works. The next lesson is that, no matter how hard you try, you always end up with a handful of leftover nuts and bolts. These remaining components can be informative: will your machine still work without them? The same logic applies to one approach to understanding human brain function: by investigating the effects of lesions in animals and accidental brain damage in humans we can ask which parts are necessary for specific functions. Over the past twenty years, it has become possible to interfere with human brain functions safely and reversibly, and to control when and where the interference is induced. The technique, known as transcranial magnetic stimulation (TMS), has become a mainstay of cognitive neuroscience.A PDF verion can be downloaded HERE.
There is controversy over the ethics of using this techniques on humans, it can cause seizures or psychosis in some subjects. This is the subject of a letter to the editor in the March 23 issue of Science from Leslie Sargent Jones of the Univ. of S. Carolina:
When Science publishes research using healthy human subjects, one assumes there was minimal risk and/or vital clinical value. This does not appear to be the case for the work by D. Knoch and colleagues ("Diminishing reciprocal fairness by disrupting the right prefrontal cortex," Reports, 3 Nov. 2006, p. 829). Their results on the dorsolateral prefrontal cortex's role in judgments of fairness and self-interest are interesting, but they largely validated what was already suspected.Experimental subjects received repetitive transcranial magnetic stimulation (rTMS) for 15 min to produce "suppression of activity in the stimulated brain region." The rTMS generated an electric maelstrom powerful enough to disrupt all activity for 7 min. Animal rTMS research (with overexposure as in LD50 drug toxicity studies) shows that anything studied (e.g., receptor levels) is modified. For rTMS in humans, known risks range from headaches to, more rarely, seizures or psychosis (1). Long-term occult changes and self-reported symptoms in healthy subjects have not been studied, and rTMS continues to be used for studies both fascinating and frivolous (just check the literature).
The use of rTMS on healthy subjects does not meet the definition of "minimal risk" (45 CFR section 46.102: risks… "not greater … than those … encountered in daily life"). We know that healthy subjects don't risk seizures or psychosis in their "daily life." What we don't know is what the residual effects of this activity-swamping tsunami of electrical current are. The Report demonstrates a naiveté about the possibility of rTMS having long-term or negative consequences. Oddly, some of these authors have used rTMS to treat neuropsychiatric disorders on the basis of its long-lasting effects (2). Roentgen's technology was also once thought harmless, and x-rays were used to check shoe sizes (3). We know better now.
- K. Machii, D. Cohen, C. Ramos-Estebanez, A. Pascual-Leone, Clin. Neurophysiol. 117, 455 (2006).
- C. M. Miller, Newsweek, "Minds and magnets," 11 Dec. 2006 (http://www.msnbc.msn.com/id/16008890/site/newsweek/).
- J. Duffin, C. R. R. Hayter, Isis 91, 260 (2000).
Thursday, March 29, 2007
Your Brain on Music
I have finally finished reading, and would like to strongly recommend, Daniel J. Levitin's book "This Is Your Brain on Music." The website for this book has fascinating musical examples. The beginning chapters of the book offer the most entertaining, efficient, and engaging explanations of music's essential elements (such as pitch, timbre, rhythym, loudness, anticipation, etc.) that I have ever seen. I also particularly liked chapters 6 and 9, on emotion and instinct.
Levitin's research has studied the involvement of more 'primitive' parts of the brain, like the cerebellum, in music comprehension and generation. In this vein the recent report by Wong et al. in Nature Neuroscience, showing that musical training can change brainstem encoding of linguistic pitch, is of interest. Here is their abstract:
Levitin's research has studied the involvement of more 'primitive' parts of the brain, like the cerebellum, in music comprehension and generation. In this vein the recent report by Wong et al. in Nature Neuroscience, showing that musical training can change brainstem encoding of linguistic pitch, is of interest. Here is their abstract:
Music and speech are very cognitively demanding auditory phenomena generally attributed to cortical rather than subcortical circuitry. We examined brainstem encoding of linguistic pitch and found that musicians show more robust and faithful encoding compared with nonmusicians. These results not only implicate a common subcortical manifestation for two presumed cortical functions, but also a possible reciprocity of corticofugal speech and music tuning, providing neurophysiological explanations for musicians' higher language-learning ability.
Interlude: some Franz Liszt
Since I am doing an adjacent post on music and the brain, I thought I would post a bit more of my playing: Les Cloches de Genève, from Années de Plèrinage, recorded last year on my Steinway B at Twin Valley, Middleton, Wisconsin.
Wednesday, March 28, 2007
Correlation of conscious perception with synchronization of neural activity across cortical areas.
Melloni et al. offer interesting measurements that correlate electrical activity with stimuli that are consciously versus unconsciously perceived. Their abstract:

Subliminal stimuli can be deeply processed and activate similar brain areas as consciously perceived stimuli. This raises the question which signatures of neural activity critically differentiate conscious from unconscious processing. Transient synchronization of neural activity has been proposed as a neural correlate of conscious perception. Here we test this proposal by comparing the electrophysiological responses related to the processing of visible and invisible words in a delayed matching to sample task. Both perceived and nonperceived words caused a similar increase of local (gamma) oscillations in the EEG, but only perceived words induced a transient long-distance synchronization of gamma oscillations across widely separated regions of the brain. After this transient period of temporal coordination, the electrographic signatures of conscious and unconscious processes continue to diverge. Only words reported as perceived induced (1) enhanced theta oscillations over frontal regions during the maintenance interval, (2) an increase of the P300 component of the event-related potential, and (3) an increase in power and phase synchrony of gamma oscillations before the anticipated presentation of the test word. We propose that the critical process mediating the access to conscious perception is the early transient global increase of phase synchrony of oscillatory activity in the gamma frequency range.

(click on image to make it larger). Scalp topography of induced gamma power and phase synchrony for the visible and invisible condition. Top row, Visible condition. Bottom row, Invisible condition. The background color indicates induced gamma power averaged in a 50–57 Hz frequency range. Each head represents the average of a 150 ms time window. Time 0 indicates the onset of the sample word. Lines connect pairs of electrodes displaying significant synchronization. The full article, which contains further color illustrations, can be downloaded HERE.
The structure of consciousness
This is the title of an essay by Buzsaki in the March 15 issue of Nature. It has the following summary of three basic architectural schemes that are present in mammalian brains:
The simplest uses strictly local wiring. In this kind of circuit, typified by the cerebellum, a few neuronal types form individual 'modules' that may be repeated as necessary. Because interaction between modules is restricted to neighbours, it is massively parallel in nature. In different species, the size of locally organized brain structures — including the basal ganglia, thalamus and cerebellum — roughly scales with the number of modules they contain.He suggests a view of subjective consciousness (that has been proposed also by several other researchers):
An entirely different type of network uses random connections, with a more or less equal probability of connecting local, intermediate or distant neurons. A unique example of such a random connectionist scheme is the recurrent excitatory circuit of the hippocampal CA3 region.
The third architectural scheme, exemplified by the neocortex, combines local modularity with more random, long-range connectivity. This complex wiring scheme shares many properties with 'small-world' or 'scale-free' networks. The advantage of this arrangement is that the number of intermediate steps between any two neurons — the synaptic path length — can remain relatively constant when network size is scaled up, because even a small fraction of long-range connections can dramatically reduce the average path length. Although intermediate and long-range interconnections demand resources and space, they are critical for globally distributing the results of local computations throughout the entire cerebral cortex.

.... that the local–global wiring of the cerebral cortex and the perpetual, self-organized complex dynamics it supports are necessary ingredients for subjective experiences. Environmental inputs can be seen as perturbations of the ongoing spontaneous activity. If they manage to perturb ongoing activity for a sufficiently long time in a big enough population of neurons, their effect will be noticed; that is, we will become conscious of them. In contrast, the locally organized cerebellar cortex, used largely for sensorimotor integration, does not give rise to self-generated or spontaneous activity, and its response to input remains local and non-persistent. Importantly, we generate no subjective record of such local computations.The complete essay can be downloaded HERE.
Tuesday, March 27, 2007
Damage to the prefrontal cortex increases utilitarian moral judgements
In other words, people with a rare injury - damage to the ventromedial prefrontal cortex - expressed increased willingness to kill or harm another person if doing so would save others’ lives. Benedict Carey's review (March 22 NY Times) of the paper by Koenigs et al. (PDF download HERE) notes that:

Lesions of the six VMPC patients displayed in mesial views and coronal slices. The colour bar indicates the number of overlapping lesions at each voxel. (Click on the figure to enlarge).
The findings are the most direct evidence that humans’ native revulsion to hurting others relies on a part of neural anatomy, one that evolved before the higher brain regions responsible for analysis and planning.Koenigs et al:
...show that six patients with focal bilateral damage to the ventromedial prefrontal cortex (VMPC), a brain region necessary for the normal generation of emotions and, in particular, social emotions, produce an abnormally 'utilitarian' pattern of judgements on moral dilemmas that pit compelling considerations of aggregate welfare against highly emotionally aversive behaviours (for example, having to sacrifice one person's life to save a number of other lives). In contrast, the VMPC patients' judgements were normal in other classes of moral dilemmas. These findings indicate that, for a selective set of moral dilemmas, the VMPC is critical for normal judgements of right and wrong. The findings support a necessary role for emotion in the generation of those judgements.

Lesions of the six VMPC patients displayed in mesial views and coronal slices. The colour bar indicates the number of overlapping lesions at each voxel. (Click on the figure to enlarge).
Innateness and culture in language evolution - a bit of heresy.
Kirby et al ask:

Fig. 1. (Click to enlarge) The structure of language arises from the interactions between three complex adaptive systems. As individuals, we acquire language using learning mechanisms that are part of our biological endowment (characterized in this paper in terms of prior bias). This learning machinery acts as the mechanism by which language is transmitted culturally through a population of individuals over time. Ultimately, this process of cultural transmission leads to a set of language universals (which can be expressed as a distribution over types of languages). The relationship between learning machinery and consequent universals is nontrivial but can be uncovered using the framework developed here. Finally, the structure of languages that emerge from this process will affect the fitness of individuals using those languages, which in turn will lead to the biological evolution of language learners, closing the loop of interactions.

Fig. 2. (Click to enlarge). The link between biological predispositions and language structure. Genes (in combination with the nonlinguistic environment) give rise to mechanisms for learning and processing language. These determine our innate predispositions with respect to language (our prior linguistic bias). Bias is a property of an individual, but the (universal) structure of human language emerges from the interaction of many individuals over time. Therefore, cultural transmission bridges the link between bias and universals. Although genes code for bias, biological fitness will in part be governed by the extended phenotype (i.e., language structure). To understand language evolution, we must understand this linking mechanism.
Although languages vary, they share many universal structural properties. Where do these universals come from? A great deal of research has proceeded under the assumption that this is essentially a biological question: that languages have the structure they do because of our innate faculty for acquiring and processing language.They suggest:
...that there are serious problems with this orthodox evolutionary/biolinguistic approach. It treats language as arising from two adaptive systems, individual learning and biological evolution, but in doing so misses a third: cultural transmission. The surprising consequences of taking all three adaptive systems into account are that strong universals need not arise from strong innate biases, that adaptation does not necessarily imply natural selection, and that cultural transmission may reduce the selection pressure on innate learning mechanisms. Our conclusions call into question the existence of strongly constraining biological predispositions for language, and the prominence of adaptationist explanations for the structural properties of languages.Here are two useful figures from the paper, and the details of the Bayesian model they use you can find in the PDF of the article.

Fig. 1. (Click to enlarge) The structure of language arises from the interactions between three complex adaptive systems. As individuals, we acquire language using learning mechanisms that are part of our biological endowment (characterized in this paper in terms of prior bias). This learning machinery acts as the mechanism by which language is transmitted culturally through a population of individuals over time. Ultimately, this process of cultural transmission leads to a set of language universals (which can be expressed as a distribution over types of languages). The relationship between learning machinery and consequent universals is nontrivial but can be uncovered using the framework developed here. Finally, the structure of languages that emerge from this process will affect the fitness of individuals using those languages, which in turn will lead to the biological evolution of language learners, closing the loop of interactions.

Fig. 2. (Click to enlarge). The link between biological predispositions and language structure. Genes (in combination with the nonlinguistic environment) give rise to mechanisms for learning and processing language. These determine our innate predispositions with respect to language (our prior linguistic bias). Bias is a property of an individual, but the (universal) structure of human language emerges from the interaction of many individuals over time. Therefore, cultural transmission bridges the link between bias and universals. Although genes code for bias, biological fitness will in part be governed by the extended phenotype (i.e., language structure). To understand language evolution, we must understand this linking mechanism.
Monday, March 26, 2007
Brain imaging can infer your hidden intentions....
As a followup to the 3/14 posting on NeuroLaw, this report from Haynes et al. seems relevant. Here is their abstract, along with a figure from their paper:
When humans are engaged in goal-related processing, activity in prefrontal cortex is increased. However, it has remained unclear whether this prefrontal activity encodes a subject's current intention. Instead, increased levels of activity could reflect preparation of motor responses, holding in mind a set of potential choices, tracking the memory of previous responses, or general processes related to establishing a new task set. Here we study subjects who freely decided which of two tasks to perform and covertly held onto an intention during a variable delay. Only after this delay did they perform the chosen task and indicate which task they had prepared. We demonstrate that during the delay, it is possible to decode from activity in medial and lateral regions of prefrontal cortex which of two tasks the subjects were covertly intending to perform. This suggests that covert goals can be represented by distributed patterns of activity in the prefrontal cortex, thereby providing a potential neural substrate for prospective memory. During task execution, most information could be decoded from a more posterior region of prefrontal cortex, suggesting that different brain regions encode goals during task preparation and task execution. Decoding of intentions was most robust from the medial prefrontal cortex, which is consistent with a specific role of this region when subjects reflect on their own mental states.Left: A spherical searchlight centered on one voxel (vi) was used to define a local neighborhood. For each scanning run, the spatial response pattern in this local spherical cluster was extracted during preparation of either subtraction or addition. We then trained a pattern classifier with a subset of the data to recognize the typical response patterns associated with covert preparation of the two mathematical operations (see Experimental Procedures) and measured the local decoding accuracy. Then, the searchlight was shifted to the next spatial location.
Middle: Highlighted in green are medial brain regions (superimposed on a saggital slice of an anatomical template image) where this local classifier was able to decode significantly above chance which intention the subjects were covertly holding in an independent test data set. Highlighted in red are regions where it was possible to decode the intention during the execution of the task.

'Thinking about thinking' shown in Rats...
A simple experiment can be used to demonstrate 'thinking about thinking', or metacognition in humans, monkeys, and bottlenose dolphins. In dealing with uncertain situations, we more likely to decline a visual, auditory or memory discrimination task that can involve reward or punishment if we are uncertain of the discrimination (more/less, same/different) being requested. Thus we are testing possible future outcomes. This behavior has not been seen in pigeons and rats.
In a recent issue of Current Biology Foote and Crystal now:
In a recent issue of Current Biology Foote and Crystal now:
demonstrate for the first time that rats are capable of metacognition—i.e., they know when they do not know the answer in a duration-discrimination test. Before taking the duration test, rats were given the opportunity to decline the test. On other trials, they were not given the option to decline the test. Accurate performance on the duration test yielded a large reward, whereas inaccurate performance resulted in no reward. Declining a test yielded a small but guaranteed reward. If rats possess knowledge regarding whether they know the answer to the test, they would be expected to decline most frequently on difficult tests and show lowest accuracy on difficult tests that cannot be declined. Our data provide evidence for both predictions and suggest that a nonprimate has knowledge of its own cognitive state.PDE of article HERE.
Subscribe to:
Posts (Atom)