I put my playing of a Debussy Reverie on YouTube about 10 months ago, and am completely amazed that it has by now had ~18,000 viewings, and over 50 largely constructive comments, many of which suggested to "slow down." So, I did a second version and posted it two months ago, which I pass on here. Response has been more favorable.
Tuesday, July 17, 2007
Genetics and tonal languages
It is likely that there are heritable differences of brain structure and function that affect language acquisition and usage...cognitive biases in a population of acquirers could influence the direction of language change across generations. These biasing effects could result in linguistic differences between populations, producing nonspurious (causal) correlations between genetic and linguistic diversities. Dediu and Ladd:
... propose that the linguistic typology of tone is affected by such a bias. Human languages differ typologically in the way they use voice fundamental frequency (pitch). All languages use consonants and vowels to distinguish one word or grammatical category from another, but, in addition, so-called "tone languages" (e.g., Chinese) use pitch for this purpose as well, whereas "non-tone languages" (e.g., English) use pitch only at sentence level (to convey emphasis, emotion, etc.). In tone languages, that is, pitch is organized into tone phonemes that are functionally comparable with consonant and vowel phonemes. Tone languages are the norm in sub-Saharan Africa and are very common in continental and insular southeast Asia. They are rare in the rest of Eurasia, North Africa, and Australia. They are relatively common in Central America, the Caribbean, and the Amazon basin, and occur sporadically elsewhere among the aboriginal languages of the Americas..Here is their Abstract:
The correlations between interpopulation genetic and linguistic diversities are mostly noncausal (spurious), being due to historical processes and geographical factors that shape them in similar ways. Studies of such correlations usually consider allele frequencies and linguistic groupings (dialects, languages, linguistic families or phyla), sometimes controlling for geographic, topographic, or ecological factors. Here, we consider the relation between allele frequencies and linguistic typological features. Specifically, we focus on the derived haplogroups (note: these are sets of nucleotide polymorphisms) of the brain growth and development-related genes ASPM and Microcephalin, which show signs of natural selection and a marked geographic structure, and on linguistic tone, the use of voice pitch to convey lexical or grammatical distinctions. We hypothesize that there is a relationship between the population frequency of these two alleles and the presence of linguistic tone and test this hypothesis relative to a large database (983 alleles and 26 linguistic features in 49 populations), showing that it is not due to the usual explanatory factors represented by geography and history. The relationship between genetic and linguistic diversity in this case may be causal: certain alleles can bias language acquisition or processing and thereby influence the trajectory of language change through iterated cultural transmission.
Monday, July 16, 2007
Most popular consciousness papers for June 2007
That is, the five most downloaded from the eprint archive maintained by the Association for the Scientific Study of Consciousness.
1. Rosen, Alan and Rosen, David B. (2006) The Design of a
Sensation-generating Mechanism in the Brain: A first step towards a
quantitative definition of consciousness. In: Consciousness and
Cognition (1223 downloads from 9 countries).
http://eprints.assc.caltech.edu/195/
2. Sagiv, Noam and Ward, Jamie (2006) Crossmodal interactions: lessons
from synesthesia. In: Visual Perception, Part 2 (1171 downloads from
20 countries). http://eprints.assc.caltech.edu/224/
3. Ruby, Perrine and Legrand, Dorothée (2007) Neuroimaging the self?
In: Sensorimotor foundations of higher cognition. OUP (882 downloads
from 14 countries). http://eprints.assc.caltech.edu/275/
4. Koriat, A. (2006) Metacognition and Consciousness. In: Cambridge
handbook of consciousness. CUP (804 downloads from 15 countries).
http://eprints.assc.caltech.edu/175/
5. Windt, Jennifer Michelle and Metzinger, Thomas (2006) The
philosophy of dreaming and self-consciousness: What happens to the
experiential subject during the dream state? In: The new science of
dreaming (780 downloads from 22 countries).
http://eprints.assc.caltech.edu/200/
1. Rosen, Alan and Rosen, David B. (2006) The Design of a
Sensation-generating Mechanism in the Brain: A first step towards a
quantitative definition of consciousness. In: Consciousness and
Cognition (1223 downloads from 9 countries).
http://eprints.assc.caltech.edu/195/
2. Sagiv, Noam and Ward, Jamie (2006) Crossmodal interactions: lessons
from synesthesia. In: Visual Perception, Part 2 (1171 downloads from
20 countries). http://eprints.assc.caltech.edu/224/
3. Ruby, Perrine and Legrand, Dorothée (2007) Neuroimaging the self?
In: Sensorimotor foundations of higher cognition. OUP (882 downloads
from 14 countries). http://eprints.assc.caltech.edu/275/
4. Koriat, A. (2006) Metacognition and Consciousness. In: Cambridge
handbook of consciousness. CUP (804 downloads from 15 countries).
http://eprints.assc.caltech.edu/175/
5. Windt, Jennifer Michelle and Metzinger, Thomas (2006) The
philosophy of dreaming and self-consciousness: What happens to the
experiential subject during the dream state? In: The new science of
dreaming (780 downloads from 22 countries).
http://eprints.assc.caltech.edu/200/
Modulating emotional appraisal by false physiological feedback.
Grey et al. examine how emotional appraisal is influenced by physiological feedback. Their observations make me wonder whether trying the opposite trick, giving false feedback that suggests less autonomic arousal, could chill out reactions to an emotional stimulus... Their main points:
James and Lange proposed that emotions are the perception of physiological reactions. Two-level theories of emotion extend this model to suggest that cognitive interpretations of physiological changes shape self-reported emotions. Correspondingly false physiological feedback of evoked or tonic bodily responses can alter emotional attributions. Moreover, anxiety states are proposed to arise from detection of mismatch between actual and anticipated states of physiological arousal. However, the neural underpinnings of these phenomena previously have not been examined.
We undertook a functional brain imaging (fMRI) experiment to investigate how both primary and second-order levels of physiological (viscerosensory) representation impact on the processing of external emotional cues. Twelve participants were scanned while judging face stimuli during both exercise and non-exercise conditions in the context of true and false auditory feedback of tonic heart rate. We observed that the perceived emotional intensity/salience of neutral faces was enhanced by false feedback of increased heart rate. Regional changes in neural activity corresponding to this behavioural interaction were observed within included right anterior insula, bilateral mid insula, and amygdala. In addition, right anterior insula activity was enhanced during by asynchronous relative to synchronous cardiac feedback even with no change in perceived or actual heart rate suggesting this region serves as a comparator to detect physiological mismatches. Finally, BOLD activity within right anterior insula and amygdala predicted the corresponding changes in perceived intensity ratings at both a group and an individual level.
Our findings identify the neural substrates supporting behavioural effects of false physiological feedback, and highlight mechanisms that underlie subjective anxiety states, including the importance of the right anterior insula in guiding second-order “cognitive” representations of bodily arousal state.
Williams Syndrome - evidence for a discrete social brain
David Dobbs offers a very well written essay (PDF here) on Williams Syndrome in the Sunday New York Times Magazine (7/8/07). The syndrome is caused by a well-defined deletion in chromosome 7 that occasionally occurs during the synthesis of egg or sperm cells. Patients have a low IQ (~60) and compromised spatial skill and analytical thought, but are hyper-sociable and friendly, very talkative. An pathway from the orbitofrontal (OFC) cortex to the amygdala that usually signals dangerous or angry faces is inactive; but curiously the OFC-amygdala connection still works normally for nonsocial threats such as pictures of snakes, sharks or car crashes. The existence of this syndrome provides perhaps the strongest evidence for genetically and developmentally distinct class of 'social brain' mechanisms distinct from other higher sensory, motor, and analytical skills.
Sunday, July 15, 2007
A Sunday Walk....
Around my house on Twin Valley Road in Town of Middleton, Wisconsin. (It is a stone schoolhouse built in 1860, converted to a residence).












Friday, July 13, 2007
How pain preempts cognition - and about itching
I found these two articles on pain and itching interesting, particularly since I've been going through an orgy of both after exposing myself to poison oak or ivy while working in the yard. Bingel et al. show some brain correlates of why I found it difficult to focus on normal cognitive activities during this period:
Now, on to itching (also central to my poison oak story, still going on....). Johanek et al. note a curious distinction between histamine induced itching and itching caused by cutaneous application of spicules from the cowhage plant. They show a different class of afferent C-fiber afferents signaling non-histamine induced itching:
It is well known that pain attracts attention and interferes with cognition. Given that the mechanisms behind this phenomenon are largely unknown, we used functional magnetic resonance imaging and presented visual objects with or without concomitant pain stimuli. To test for the specificity of pain, we compared this modulatory effect with a previously established modulatory effect of working memory on visual object processing. Our data showed a comparable behavioral effect of both types of modulation and identified the lateral occipital complex (LOC) as the site of modulation in the ventral visual stream, for both pain and working memory. However, the sources of these modulatory effects differed for the two processes. Whereas the source of modulation for working memory could be attributed to the parietal cortex, the modulatory effect of pain was observed in the rostral anterior cingulate cortex (rACC), an area ideally suited to link pain perception and attentional control.A) fMRI effects of the interaction of background visibility with working memory load were observed in bilateral LOC, reflecting a phasic modulation of LOC activity.
(B) Corresponding activation and parameter estimates related to the interaction of Pain × Visibility.

Now, on to itching (also central to my poison oak story, still going on....). Johanek et al. note a curious distinction between histamine induced itching and itching caused by cutaneous application of spicules from the cowhage plant. They show a different class of afferent C-fiber afferents signaling non-histamine induced itching:
The neuronal pathways for itch have been characterized mainly based on responses to histamine. Intracutaneous application of histamine produces intense itch and a large area of axon-reflexive vasodilation ("flare") around the application site. Both phenomena are thought to be mediated through neuronal activity in itch-specific, mechanoinsensitive C-fiber afferents (CMi). However, mechanical and electrical stimuli that do not activate CMi fibers can cause the sensation of itch, and itch may occur without flare, suggesting that other neuronal itch pathways exist. Because cutaneous application of spicules from the plant Mucuna pruriens (cowhage) has been anecdotally reported to produce itch without flare, we performed psychophysical experiments to investigate whether the mechanisms underlying cowhage- and histamine-induced itch differ. Although histamine and cowhage produced itch of similar magnitude, the itch to cowhage was not correlated with the itch to histamine; some subjects had intense itch to cowhage and little itch to histamine and visa versa. Laser Doppler measurements of blood flow revealed that histamine led to a large area of vasodilation, whereas cowhage produced vasodilation restricted to the application site. Pretreatment of the skin with an antihistamine blocked the itch produced by histamine but did not prevent cowhage-induced itch. Desensitization of the skin with topical capsaicin abolished cowhage-induced itch but did not significantly alter histamine-induced itch. These findings indicate that cowhage itch is signaled through a population of capsaicin-sensitive afferent nerve fibers that is distinct from CMi fibers mediating histamine-induced itch. Cowhage may be useful to investigate the neural pathway mediating nonhistaminergic itch.
Negative suggestions enhancing pain -an antidote?
Benedetti et al. provide an interesting review of the nocebo effect (the anticipation of pain enhancing its magnitude). This is the opposite of the placebo effect, where a more positive expectation can lower perceived pain. Their article (PDF here) provides some interesting graphics of brain imagining showing this effect, and discusses relevant brain receptors.
Recent experimental evidence indicates that negative verbal suggestions induce anticipatory anxiety about the impending pain increase, and this verbally-induced anxiety triggers the activation of cholecystokinin (CCK) which, in turn, facilitates pain transmission. CCK-antagonists have been found to block this anxiety-induced hyperalgesia, thus opening up the possibility of new therapeutic strategies whenever pain has an important anxiety component.

Recent experimental evidence indicates that negative verbal suggestions induce anticipatory anxiety about the impending pain increase, and this verbally-induced anxiety triggers the activation of cholecystokinin (CCK) which, in turn, facilitates pain transmission. CCK-antagonists have been found to block this anxiety-induced hyperalgesia, thus opening up the possibility of new therapeutic strategies whenever pain has an important anxiety component.

Thursday, July 12, 2007
Rodent Altruism - Lending an Anonymous Paw
Krista Zala writes a nice summary of work by Rutte and Taborsky. Some clips:
Rats given a helping paw are more prone to helping others--even complete strangers. This suggests that the animals' social life may be richer than we thought...Many animals, including rats, demonstrate direct reciprocity--described as "I'll help you if you help me." But generalized reciprocity, in which individuals remember how they were treated in the recent past and apply it to others, including strangers, has been thought to be a uniquely human trait...However, previous studies haven't specifically looked for generalized reciprocity in other animals.The New York Times Science section has now also done a popular summary of this work (PDF here), and the graphic above is from that review.
To find out if rats have this capacity, Rutte and Michael Taborsky of the University of Bern in Switzerland trained rats to pull a lever that would deliver an oat flake reward to another rat on the other side of a wire mesh wall in a shared cage. Some of these rats were then put on the receiving side, paired for several days with either other rats trained to be helpful—-three different ones over the span--or with untrained rats that didn't pull the lever and provide food. After several days of living with such generous or not-so-generous neighbors, these test rats were then switched back to the lever side of the cage, paired with a new neighbor rat, and watched to see if they would provide food for it. Rats who had been paired with food-providing neighbors helped their new partner more often than those who had had unhelpful neighbors. Rutte's team also found that when a test rat was paired with one of the rats that had earlier provided it with oak flakes, it pulled the food lever even more--showing direct reciprocity. When the cage was empty of any neighbor rat, it barely pulled the food lever at all.
In a pack of 200 rats, where it's hard to remember who's been helpful and who hasn't, a general willingness to help others makes sense as a strategy...It may be a mechanism for how cooperation can evolve when you cannot recognize your partner...rats are notoriously bad at remembering other individuals.
Looking into the prevalence of generalized reciprocity among social and nonsocial animals may help our understanding of the evolution of cooperation.

Altruistic Chimps and Children
As a companion to the blog posting on helping rodents, this work from Tomasello's laboratory. The article has some nice videos. Here is the author's summary:
Debates about altruism are often based on the assumption that it is either unique to humans or else the human version differs from that of other animals in important ways. Thus, only humans are supposed to act on behalf of others, even toward genetically unrelated individuals, without personal gain, at a cost to themselves. Studies investigating such behaviors in nonhuman primates, especially our close relative the chimpanzee, form an important contribution to this debate. Here we present experimental evidence that chimpanzees act altruistically toward genetically unrelated conspecifics. In addition, in two comparative experiments, we found that both chimpanzees and human infants helped altruistically, regardless of any expectation of reward, even when some effort was required, and even when the recipient was an unfamiliar individual—all features previously thought to be unique to humans. The evolutionary roots of human altruism may thus go deeper than previously thought, reaching as far back as the last common ancestor of humans and chimpanzees.
Wednesday, July 11, 2007
Neuroimaging of emotions driving political strategy...
My October 23 post mentioned the work of Westen, who has used functional neuroimaging to show how emotions prevail over reason when people rationalize their political choices. This material - up to date neuroscience of decision making - has been taken up and absorbed as a guide to campaign strategy in an amazingly short time, with democratic party strategists particularly engaged by Westen's recent book, “The Political Brain: The Role of Emotion in Deciding the Fate of the Nation” (Public Affairs).
Westen takes the unlikely position that the Democratic Party should, for the most part, forget about issues, policies, even facts, and instead focus on feelings. His book takes a very different tack than, say, “What’s the Matter With Kansas?” by Thomas Frank or Al Gore’s “Assault on Reason,” which try to explain voter behavior in terms of self-interest and factual analysis.
The recent review of Westen's book by Patricia Cohen in the New York Times, notes that Westen has by now given numerous talks to political groups and set up a political consulting company. She notes Bill Clinton's positive reaction to the book, as he "particularly liked the discussion of how one could evoke emotion without being intellectually dishonest.” Cohen's review is worth reading (PDF here).
In this vein, it is interesting that Louis Menand in a recent New Yorker review titled "The Fractured Franchise" has an elegant discussion of evidence that most voters are massively ignorant and misinformed about political reality (PDF here). Thus, even when 'democracy works' the results can be a disaster. (So we should prefer rule by economists?) He reviews the recent book by Bryan Caplan, “The Myth of the Rational Voter: Why Democracies Choose Bad Politics” (Princeton; $29.95). (By the way, see also Menand's earlier article "The Unpolitical Animal." He is a brilliant guy. )
Westen takes the unlikely position that the Democratic Party should, for the most part, forget about issues, policies, even facts, and instead focus on feelings. His book takes a very different tack than, say, “What’s the Matter With Kansas?” by Thomas Frank or Al Gore’s “Assault on Reason,” which try to explain voter behavior in terms of self-interest and factual analysis.
The recent review of Westen's book by Patricia Cohen in the New York Times, notes that Westen has by now given numerous talks to political groups and set up a political consulting company. She notes Bill Clinton's positive reaction to the book, as he "particularly liked the discussion of how one could evoke emotion without being intellectually dishonest.” Cohen's review is worth reading (PDF here).
In this vein, it is interesting that Louis Menand in a recent New Yorker review titled "The Fractured Franchise" has an elegant discussion of evidence that most voters are massively ignorant and misinformed about political reality (PDF here). Thus, even when 'democracy works' the results can be a disaster. (So we should prefer rule by economists?) He reviews the recent book by Bryan Caplan, “The Myth of the Rational Voter: Why Democracies Choose Bad Politics” (Princeton; $29.95). (By the way, see also Menand's earlier article "The Unpolitical Animal." He is a brilliant guy. )
Hi-Tech fMRI lie detection - another scam?
Margaret Talbot writes an interesting article in The New Yorker (PDF here) about the current hype over lie detection using brain scanning techniques, and the sprouting of numerous companies wanting to sell their services to private investigators, police departments, U.S. and foreign government agencies, etc. The article is a broad discussion of lie detection also by other physiological indicators such as skin conductance and blood pressure. Most interesting to me were the critical points raised. Here are a few clips from the article:
Paul Bloom, a cognitive psychologist at Yale, believes that brain imaging has a beguiling appeal beyond its actual power to explain mental and emotional states. "Psychologists can be heard grousing that the only way to publish in Science or Nature is with pretty color pictures of the brain," he wrote in an essay for the magazine Seed. "Critical funding decisions, precious column inches, tenure posts, science credibility, and the popular imagination have all been influenced by fMRI's seductive but deceptive grasp on our attentions." Indeed, in the past decade, Nature alone has published nearly a hundred articles involving fMRI scans. The technology is a remarkable tool for exploring the brain, and may one day help scientists understand much more about cognition and emotion. But enthusiasm for brain scans leads people to overestimate the accuracy with which they can pinpoint the sources of complex things like love or altruism, let alone explain them.Brain scans enthrall us, in part, because they seem more like "real" science than those elaborate deductive experiments that so many psychologists perform. In the same way that an X-ray confirms a bone fissure, a brain scan seems to offer an objective measure of mental activity. And, as Bloom writes, fMRI research "has all the trappings of work with great lab-cred: big, expensive, and potentially dangerous machines, hospitals and medical centers, and a lot of people in white coats."
Deena Skolnick Weisberg, a graduate student at Yale, has conducted a clever study, to be published in the Journal of Cognitive Neuroscience, which points to the outsized glamour of brain-scan research. She and her colleagues provided three groups--neuroscientists, neuroscience students, and ordinary adults--with explanations for common psychological phenomena (such as the tendency to assume that other people know the same things we do). Some of these explanations were crafted to be bad. Weisberg found that all three groups were adept at identifying the bad explanations, except when she inserted the words "Brain scans indicate." Then the students and the regular adults became notably less discerning. Weisberg and her colleagues conclude, "People seem all too ready to accept explanations that allude to neuroscience."
Nancy Kanwisher, a cognitive scientist at M.I.T., relies a great deal on MRI technology. In 1997, she identified an area near the bottom of the brain that is specifically involved in perceiving faces. She has become a pointed critic of the rush to commercialize brain imaging for lie detection, and believes that it's an exaggeration even to say that research into the subject is "preliminary." The tests that have been done, she argues, don't really look at lying. "Making a false response when instructed to do so is not a lie," she says. The ninety-per-cent "accuracy" ascribed to fMRI lie detection refers to a scenario so artificial that it is nearly meaningless. To know whether the technology works, she believes, "you'd have to test it on people whose guilt or innocence hasn't yet been determined, who believe the scan will reveal their guilt or innocence, and whose guilt or innocence can be established by other means afterward." In other words, you'd have to run a legal version of a clinical trial, using real suspects instead of volunteers.
She points out that the various brain regions that appear to be significantly active during lying are "famous for being activated in a wide range of different conditions--for almost any cognitive task that is more difficult than an easier task." She therefore believes that fMRI lie detection would be vulnerable to countermeasures--performing arithmetic in your head, reciting poetry--that involve concerted cognitive effort. Moreover, the regions that allegedly make up the brain's "lying module" aren't that small...Saying 'You have activation in the anterior cingulate' is like saying 'You have activation in Massachusetts.' "
Kanwisher's complaint suggests that fMRI technology, when used cavalierly, harks back to two pseudosciences of the eighteenth and nineteenth centuries: physiognomy and phrenology. Physiognomy held that a person's character was manifest in his facial features; phrenology held that truth lay in the bumps on one's skull. In 1807, Hegel published a critique of physiognomy and phrenology in "The Phenomenology of Spirit." In that work, as the philosopher Alasdair MacIntyre writes, Hegel observes that "the rules that we use in everyday life in interpreting facial expression are highly fallible." (A friend who frowns throughout your piano recital might explain that he was actually fuming over an argument with his wife.) Much of what Hegel had to say about physiognomy applies to modern attempts at mind reading.
Elizabeth Phelps, a prominent cognitive neuroscientist at N.Y.U., who studies emotion and the brain, questions another basic assumption behind all lie-detection schemes--that telling a falsehood creates conflict within the liar. With the polygraph, the assumption is that the conflict is emotional: the liar feels guilty or anxious, and these feelings produce a measurable physiological response. With brain imaging, the assumption is that the conflict is cognitive:the liar has to work a little harder to make up a story, or even to stop himself from telling the truth. Neither is necessarily right. "Sociopaths don't feel the same conflict when they lie," Phelps says. "The regions of the brain that might be involved if you have to inhibit a response may not be the same when you're a sociopath, or autistic, or maybe just strange. Whether it's an emotional or a cognitive conflict you're supposed to be exhibiting, there's no reason to assume that your response wouldn't vary depending on what your personal tendencies are--on who you are."

Tuesday, July 10, 2007
When watching a tactile stroke is the same as receiving one.
An interesting report from Banissy and Ward (PDF here)... People who have 'mirror-touch' synesthesia, when watching another person being touched, have the same experience as being touched themselves...an extreme form of empathy! They developed a protocol to provide evidence for the authenticity of this form of synesthesia, in which participants have difficulty in discriminating between actual and synesthetic touch. Their abstract:
Watching another person being touched activates a similar neural circuit to actual touch and, for some people with 'mirror-touch' synesthesia, can produce a felt tactile sensation on their own body. In this study, we provide evidence for the existence of this type of synesthesia and show that it correlates with heightened empathic ability. This is consistent with the notion that we empathize with others through a process of simulation.
Making up our Minds...
Rebecca Saxe's review of "Making Up the Mind" by Chris Frith (Blackwell, 2007, 232 pp, paperback, $24.99) is a very nice summary of why we need an empirical science of the mind that does not rely on introspection, noting that experiences of how minds work, both our own and other people's, are just fantasies whose predictions often coincide with reality. Frith's book argues that there is no qualitative difference between perceiving the outside world and our own actions in it, compared with trying to infer the thoughts, feelings and goals inside other people's heads. Here is Saxe's review:
Perceiving the outside world and one's own actions in it might seem easy, at least when compared with trying to infer the thoughts, feelings and goals inside other people's heads. These intuitions are wrong, however, and debunking them is the main task of Chris Frith's new book, Making Up the Mind. The apparently effortless perceptions of the world and the self mask sophisticated computation and 'unconscious inferences'. On the other hand, he claims, these same kinds of inferences support access to other people's feelings and intentions. There is no qualitative difference between perceiving minds and perceiving anything else.
If perceiving the world is a hard problem, studying these perceptions is possibly a harder one. By their nature, 'unconscious inferences' are not accessible to introspection. Indeed, the major trick of the scientific psychologist's trade is to devise alternative nonintrospective methods for studying the mind. The need for such methods arises not just because conscious experiences are 'soft', imprecise or hard to verify, the reasons that Frith entertains in his prologue. More importantly, as he shows in the rest of the book, the contents of introspection are simply unreliable as guides to the real structure of the mind.
Frith is especially enthusiastic about one such method: studying the brain. Brain function provides a window onto the intermediate stages of mental computations, between the inputs at the senses and the outputs in behavior and conscious experience. These days, this approach is called 'cognitive neuroscience' and its popularity is booming with the increasing availability of noninvasive brain scanners.
Making Up the Mind is an accessible and enthusiastic introduction to the field, and a great way to whet the appetites of smart beginners. Particularly nice is Frith's clear affection for doing science. During one experiment, he remembers, "we all had a very exciting Saturday in the imaging laboratory, which is not reflected at all in the paper we wrote about it afterwards." At another point, he expands on a reference to "my lab" with this footnote, "In the 1960s, this was a small bathroom that had been converted into a 'laboratory' by putting a sheet of hardboard over the bathtub."
Part 1 of the book covers the most charismatic discoveries of cognitive and neuropsychology, the gaps between the real world and the world constructed by the mind: phantom limbs, blindsight, anosognosia, change blindness, color constancy (including a fantastic demo in the color plates), priming, motor adaptation, illusions of causal control, synesthesia, implicit memory, dreams and hallucinations. (Surprisingly, he doesn't mention confabulation or false memories, two other famous illustrations of his thesis that our brains 'make up' our world.) The message is that nothing that appears simple or direct in our perceptions of the world, and of ourselves, can be taken for granted.
So how does the brain construct the consciously experienced world? No one knows. Instead of giving an answer, Part 2 introduces some basic concepts: neurons and synapses, associative and operant conditioning, Bayesian inference, and forward and inverse models. The result is a useful informal survey of the foundations of current research.
Finally, in Part 3, Frith turns back to problems of perception, now focusing on the question of social perception: how we can ever 'know' about someone else's thoughts, feelings or goals? The situation, Frith claims, is not as bad as it looks. Other people's actions are represented in the same way we represent our own actions, using a forward model to predict their causal effects. Frith illustrates this idea with an illusion. When a sound acts as a signal to start an action, the cause (the sound) and the effect (the hand motion) are perceived to be closer together in time than they really are, both for one's own and other people's actions, but not for an externally caused event. Another great example that Frith does not mention, which was reported by Sebanz and colleagues, is that the (task-irrelevant) direction that an arrow is pointing interferes with participants' ability to respond on the basis of color, if and only if the arrow points to another possible action, either for the participant or for another person. That is, a representation of another person's possible (but unseen) actions can compete with the participant's own action plans.
In all, Making Up the Mind is an accessible, fun and up-to-date introduction to the hot ideas and phenomena in and around cognitive neuroscience. Only the conclusion feels out of place. In it, Frith writes that we all do successfully 'read minds', achieving direct access to other people's thoughts and goals. How we take this conclusion depends on what Frith means by success. It is true that inferences about other minds are not qualitatively harder than other inferences, but neither are they (as Frith sometimes implies) any easier or more accurate. Inferences about other minds have the same structure as self-perception, going beyond the data given using a sophisticated combination of prior knowledge and current data, and then masked by an illusion of effortlessness. As a result, inferences about others are just as prone to leaps, gaps and confabulations. The conclusion ought to leave us where the book began in the prologue. Psychologists who want to study the mind scientifically do not rely on social perceptions of the participants any more than they rely on introspection, precisely because social mind-reading fails. Our experiences of how minds work, both our own and other people's, are just fantasies whose predictions often coincide with reality. That is why we still do need an empirical science of the mind.
Monday, July 09, 2007
Tone deafness is associated with deficits in spatial processing
Representations of pitch and space in the brain seem to interact. An article from Douglas & Bilkey in Nature Neuroscience (PDF here) now reports that people with amusia, a pitch processing deficit, do not show the interference between these concepts that is found in control subjects. The abstract:

Amusia (commonly referred to as tone-deafness) is a difficulty in discriminating pitch changes in melodies that affects around 4% of the human population. Amusia cannot be explained as a simple sensory impairment. Here we show that amusia is strongly related to a deficit in spatial processing in adults. Compared to two matched control groups (musicians and non-musicians), participants in the amusic group were significantly impaired on a visually presented mental rotation task. Amusic subjects were also less prone to interference in a spatial stimulus-response incompatibility task and performed significantly faster than controls in an interference task in which they were required to make simple pitch discriminations while concurrently performing a mental rotation task. This indicates that the processing of pitch in music normally depends on the cognitive mechanisms that are used to process spatial representations in other modalities.Here is a graphic from the review of this work by Janata in the same issue of Nature Neuroscience.

Figure... The link between musical and spatial processing was investigated via a set of tasks. (a) In a stimulus-response compatibility task, subjects press either the closer or farther of two buttons on a computer keyboard to indicate whether the second of two pitches is higher or lower than the first. The response that the second pitch was higher was made more quickly, on average, when the answer "higher pitch" was mapped to the 'higher' (farther) of the two response buttons, than when the response "lower pitch" was mapped to the higher button. (b) In the contour violation task from the Montreal Battery for the Evaluation of Amusia, subjects have to detect whether a single note in the repetition of a melody changed direction. Amusic individuals have extraordinary difficulty with this task. (c) In the Shepard and Metzler mental rotation task, subjects must determine whether two geometric figures are the same or different. (d) In the animal matching task, a large set of 15 animal pictures is shown to the subject along with a set of three probe pictures. Subjects must determine whether all three pictures from the small set are in the large set.
Ambiguity and anxiety: overreaction and a serotonin receptor
Nader and Alleine describe work of Tsetsenis et al.
Here is the abstract from Tsetsenis et al.
When the threat of terrorist attack is elevated, the United States Department of Homeland Security changes its prediction of danger from yellow to orange to red. Most of us can manage our levels of vigilance and of anxiety appropriately in response to these cues. However, imagine how debilitating it would be if you were unable to manage your anxiety and reduce your fear of attack when the threat level was reduced. In fact, many people with anxiety disorders suffer from precisely this kind of condition. The paper by Tsetsenis et al. finds that mice lacking the serotonin 1a receptor overreact to ambiguous predictors of aversive events in this same way, providing insight into factors that could predispose individuals to such disorders and into the neural locus of the effect.
Understanding the neural bases of contingency is not simply an academic question, but very much a mental health one. Unpredictable aversive events can be much more stressful than the same events when they are anticipated. In addition, psychopathologies can influence the perception of contingency. For example, depressed people have a different sense than nondepressed people of how their responses affect the environment, and exposure to unpredictable or uncontrollable aversive events is suggested to directly influence the development of depression.
The new study by Tsetsenis et al. makes a significant contribution to our understanding of the neural mechanisms mediating contingency learning. The authors studied mice in which the serotonin 1a receptor (Ht1a) gene was knocked out or inactivated during development. This receptor causes membrane hyperpolarization of nonserotonergic neurons and acts as an autoreceptor on serotonergic neurons in the raphe. Ht1a dysfunction is linked to anxiety disorders and depression, and mice lacking Ht1a receptors show increased avoidance behavior. This phenotype is attributable to the absence of the Ht1a in the forebrain during development; eliminating these receptors during adulthood does not cause the mice to show the anxious phenotype. Although fear of an aversive context is comparable in knockout and wild-type mice, the knockout mice over-generalize their fear of the 'aversive' context to a similar context containing novel elements, a situation in which wild-type mice are able to decrease their fear levels11. This finding suggested that these mutant mice focus unduly on cues that have been paired with shock rather than on cues that have not.Figure: Humans and other animals can accurately estimate the probability of danger from their experience of specific environments or cues and use this information to respond appropriately. A normal mouse (top) accurately estimates the threat from an ambiguous cue, a sleeping cat, and a less ambiguous cue, an alert cat, and is appropriately cautious or alarmed, respectively. In contrast, anxious people and animals, such as the Htr1a knockout mouse assessed by Tsetsenis et al. (bottom), often overestimate the danger represented by ambiguous cues and over-respond, given the level of threat. This is likely to interfere with the need to respond to other important events in the environment (such as cheese).

Here is the abstract from Tsetsenis et al.
Serotonin receptor 1A knockout (Htr1aKO) mice show increased anxiety-related behavior in tests measuring innate avoidance. Here we demonstrate that Htr1aKO mice show enhanced fear conditioning to ambiguous conditioned stimuli, a hallmark of human anxiety. To examine the involvement of specific forebrain circuits in this phenotype, we developed a pharmacogenetic technique for the rapid tissue- and cell type–specific silencing of neural activity in vivo. Inhibition of neurons in the central nucleus of the amygdala suppressed conditioned responses to both ambiguous and nonambiguous cues. In contrast, inhibition of hippocampal dentate gyrus granule cells selectively suppressed conditioned responses to ambiguous cues and reversed the knockout phenotype. These data demonstrate that Htr1aKO mice have a bias in the processing of threatening cues that is moderated by hippocampal mossy-fiber circuits, and suggest that the hippocampus is important in the response to ambiguous aversive stimuli.
Friday, July 06, 2007
Threads of our lives in dreams...
 Rebecca Cathcart writes an article on dreaming and "big dreams" (PDF here) that resonates with my own experience of having, particularly before waking, emotionally intense dreams whose story line seems to be an obvious attempt to integrate important personal issues. Some clips:
Rebecca Cathcart writes an article on dreaming and "big dreams" (PDF here) that resonates with my own experience of having, particularly before waking, emotionally intense dreams whose story line seems to be an obvious attempt to integrate important personal issues. Some clips:Big dreams are once again on the minds of psychologists as part of a larger trend toward studying dreams as meaningful representations of our concerns and emotions...The dreaming imagination does not just harvest images from remembered experience...It has a “poetic creativity” that connects the dots and “deforms the given,” turning scattered memories and emotions into vivid, experiential vignettes that can help us to reflect on our lives....Cultural narratives in regions like Vietnam and North and South America assign special importance to such dreams and consider them actual encounters with the spirits of lost loved ones...This notion is so widely shared by traditions all across the globe that some scholars have gone so far as to argue that religion itself actually originated in dream experience.The article emphasizes the role of dreams in dealing with death and grief.
Grief itself is transformative. It is a process of disassembly. The bereaved must let go of the selves they were, as well as the loved ones they have lost. The dreams we have while grieving are an important part of that process...Our dreams have to do with how we internalize the people we love...You learn to look within for the loved one and the particular function that person played in your life, such as caretaking or guidance in the case of a parent. This becomes part of a function that you can provide for yourself.
Dreams that occur during rapid eye movement, or REM, cycles are the most memorable and emotionally powerful...The dreams have power because brain activity during REM is most similar to that of a waking state. The emotional responses to REM dream content, therefore, are most like the responses during waking cognition...Core body temperature rises gradually from its nadir in the middle of the night during slow-wave sleep, the least active brain state. As morning nears, subcortical brain activity tied to the circadian cycle increases. When these cycles coincide in the last and longest REM phase... the mind produces its most dramatic dreams...the four or five phases of REM in a normal night’s sleep might include similar dream content. Just as the image of a lost loved one stimulates parts of the brain associated with loss, the content of dreams early in the sleep cycle could set the tone for that night’s dream experiences. Our memories upon waking, therefore, may be our recollection of a night’s cumulative dream content.
Thursday, July 05, 2007
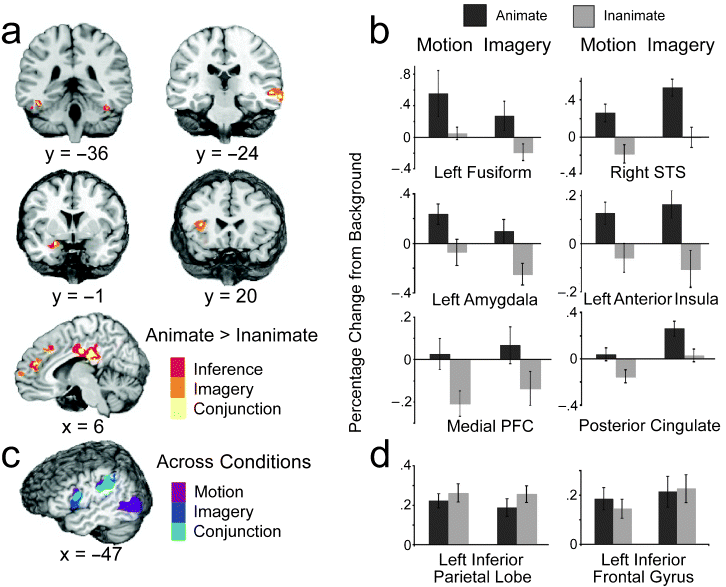
Where the brain understands animate agents..
Wheatley et al offer an interesting study in a recent issue of Psychological Science (vol 18, pg 469, 2007, PDF here). Here is the abstract and two figures:
How people understand the actions of animate agents has been vigorously debated. This debate has centered on two hypotheses focused on anatomically distinct neural substrates: The mirror-system hypothesis proposes that the understanding of others is achieved via action simulation, and the social-network hypothesis proposes that such understanding is achieved via the integration of critical biological properties (e.g., faces, affect). In this study, we assessed the areas of the brain that were engaged when people interpreted and imagined moving shapes as animate or inanimate. Although observing and imagining the moving shapes engaged the mirror system, only activation of the social network was modulated by animacy.Lateral and medial views of the social network (top, highlighted in yellow) and mirror system (bottom, highlighted in blue). The social network includes areas associated with biological motion (superior temporal sulcus, labeled "1"), biological form (lateral fusiform gyrus, labeled "6"), mentalizing (medial prefrontal cortex and posterior cingulate, labeled "3" and "4," respectively), and affective processing (insula and amygdala, labeled "2" and "5," respectively). The mirror system consists of the inferior parietal cortex (labeled "7") and the ventral-premotor/inferior-frontal cortex (labeled "8").
Experimental results. The brain slices in (a) depict areas of the social network that were more active when moving shapes were inferred (red) or imagined (orange) as animate than when they were inferred or imagined as inanimate. Yellow areas were more active for both animate inference and imagery ("conjunction"). The graph in (b) displays the average hemodynamic responses within the conjunction areas as a function of animacy (animate, inanimate) and condition (motion, imagery). (Results are not shown for the posterior insula, although this was also a conjunction area.) The illustration in (c) shows areas of the mirror system that were more active when subjects watched and made inferences about the moving shapes (purple) and when they imagined (dark blue) the moving shapes relative to when they viewed the backgrounds alone; light-blue areas were more active during both the motion and imagery conditions ("conjunction") than in the background condition. The graph in (d) shows the average hemodynamic responses of the conjunction mirror areas as a function of animacy and condition. For purposes of illustration, all group data are presented on the N27 (AFNI software) brain. Error bars represent standard errors. STS = superior temporal sulcus; PFC = prefrontal cortex.

Yawn to cool your brain?
 A curious and slightly flakey bit: Eric Nagourney in Tuesday's Science section of the NY Times describes work by Gallup et al (PDF here) published in the Journal Evolutionary Psychology. It seems to me they might have actually measured brain temperature instead of just speculating about it. Nagourney notes the proposal by Gallup et al. that:
A curious and slightly flakey bit: Eric Nagourney in Tuesday's Science section of the NY Times describes work by Gallup et al (PDF here) published in the Journal Evolutionary Psychology. It seems to me they might have actually measured brain temperature instead of just speculating about it. Nagourney notes the proposal by Gallup et al. that:yawning... is a way for the body to cool the brain...volunteers yawned more often in situations in which their brains were likely to be warmer...To prove their theory that yawning regulates brain temperature when other systems in the body are not doing enough, the researchers took advantage of the well-established tendency of people to yawn when those around them do — the so-called contagious yawn...The volunteers were asked to step into a room by themselves and watch a video showing people behaving neutrally, laughing or yawning. Observers watching through a one-way mirror counted how many times the volunteers yawned...Some volunteers were asked to breathe only through their noses as they watched. Later, volunteers were asked to press warm or cold packs on their foreheads...“The two conditions thought to promote brain cooling (nasal breathing and forehead cooling) practically eliminated contagious yawning,” the researchers wrote.
The study may also help explain why yawning spreads from person to person...A cooler brain, Dr. Gallup said, is a clearer brain...So yawning actually appears to be a way to stay more alert. And contagious yawning, he said, may have evolved to help groups remain vigilant against danger.
Subscribe to:
Posts (Atom)